ПРЕДИСЛОВИЕ
В связи с реализацией Программы форсированного инновационного индустриального развития (ФИИР) в Казахстане производство отдельных видов экологически чистых продуктов питания ставится на промышленную основу, что в определенной степени влияет на организм животного, так как сопровождается высокой плотностью животных в условиях крупногруппового содержания, связанной с гиподинамией. При этих условиях на первый план выходят необходимость профессионального подхода по профилактике заболевании животных, не допущения снижения их продуктивности, воспроизводительных функции, предотвратить другие негативные последствия.
Существующие промышленные животноводческие комплексы специализируются по выращиванию крупного рогатого скота, свиней, мелкого рогатого скота и других видов животных. Эффективность увеличения животноводческой базы за счет промышленных животноводческих комплексов несомненна. Но животные, выращенные в условиях комплексов, имеют специфические особенности, связанные с условиями содержания, кормления, селекции и гибридизации, которые отрицательно могут влиять на качественные показатели готового продукта. Наряду с положительными результатами (интенсивность выращивания, повышение мясности либо молочной продукции гарантированного качества) животные, выращенные в комплексах, приобретают ряд особенностей, которые в определенной степени будут влиять на дальнейшую технологическую обработку.
Животноводческий комплекс промышленного типа – это, крупное специализированное производство, где на основе техники и научной организации труда осуществляются непрерывные технологические процессы, обеспечивающие ритмичный выпуск продукции. Условия содержания, кормления, цикличность, зоотехнических операций, использование генетического разнообразия пород в промышленных комплексах предопределяют интенсивное выращивание скота. Поэтому в современных животноводческих комплексах используют лучшие мировые генетические ресурсы пород скота, новые методы селекции, создание на основе скрещивания и гибридизации синтетических генотипов, которые должны обеспечить устойчивый прогресс и повышение мясной и молочной продуктивности. Разрабатываются новые, более совершенные системы нормирования кормления и оценки питательности кормов.
Общеизвестно, что в условиях повышенной концентрации животных, значительный стресс в первую очередь воздействует на нервно-гуморальную и опорно-двигательную систему организма животных. Сосудистая система выполняет ряд жизненных функций в организме: регулирует кровоток, терморегуляцию, выполняет защитную функцию, доставляет к клеткам, тканям и органам питательные вещества, кислород и выносит от них продукты метаболизма, принимает непосредственное участие в создании внутренней среды (гомеостаза) организма, а вены активно реагируют на различные динамические нагрузки. Установлено, что в отдельные периоды жизни животного, в зависимости от интенсивности роста и развития органов, частей тела, имеют место характерные отличия в обменных процессах и неодинаковом использовании питательных веществ. Всё это отражается на росте, развитии животных и на их сердечнососудистой системе. Функциональные взаимоотношения с соответствующими органами и системами организма животного, особенно, динамические нагрузки, в первую очередь, влияют на рост, развитие сосудов и на аппарат конечностей.
У взрослого животного приток крови к сердцу по венам и отток от него по артериям - строго сбалансирован. В развивающемся организме, особенно у предпдода, плода, новорожденного, молодняка, наоборот, приток крови по артериям на периферию организма превалирует над её оттоком по венам с периферии к сердцу. Только в этом случае, указывают авторы, возможно создание «поля роста» для деления молодых клеток, образование тканей, формирование из зачатков органов, систем, линейного роста и увеличение массы животного.
Опорно-двигательный аппарат копытных сформировался под действием динамических нагрузок. При гиподинамии он больше испытывает статические и меньше динамические нагрузки, особенно, стопа передней и задней конечности в условиях промышленных комплексов. Появляются различного рода заболевания копытец, чаще задней конечности. Учитывая, что на изменение функциональной нагрузки первой реагирует костная ткань и её сосуды, авторы исследовали структуру компактного костного вещества трубчатых костей свободных отделов скелета конечностей, фаланг пальцев и их сосуды, суставной хрящ при дозированном принудительном движении (ДПД) и гиподинамии.
Установлено, что содержание животных при различной степени ДПД отражается на строении костей, толщине суставных хрящей, магистральных и внутриорганных костных вен, и в целом на структуре костей фаланг пальцев. При гиподинамии понижается прочность кости третьей фаланги, происходит запустевание и облитерация внутрикостных вен, понижается их клапанный индекс, что приводит к развитию болезней копытцев. При ДПД, наоборот, повышается прочность кости, особенно III фаланги, клапанный индекс, увеличивается просвет вен, толщина их стенок, понижается частота заболеваний конечностей.
Предложенная доза ДПД на расстоянии 3,5–4,0 км/час животным, выращиваемых в промышленных комплексах, является оптимальной и способствует профилактике заболеваний конечностей, увеличивает энергию роста и развития животных, повышает мясную продуктивность и качество мяса.
Выявленные особенности топографии, роста сосудов, анастомозы и коллатерали артерий вен помогут ветеринарным врачам-практикам раскрыть этиологию и установить патогенез заболеваний конечностей, в т.ч. копытец, а при оперативных вмешательствах и разработке экспериментальных приёмов - введение в венозное русло стопы инъекций крови и лекарственных препаратов.
Впервые, исходя из экспериментального материала и собственного опыта, авторами подготовлен к изданию великолепно проиллюстрированное учебное пособие, которое поможет студентам вузов, специалистам в области зоогигиены, экологии и животноводства разобраться и устранить из практики промышленных комплексов гипотрофизм и объяснить патогенез многих заболеваний животных, связанный с ним.
Капитальный научный труд российско-казахстанских ученых по изучению кровеносных сосудов и развитие периферического скелета в условиях гипо- и гипердинамии, восполнил значительный пробел по столь актуальной проблеме промышленных мясомолочных комплексов производящих экологически чистую продукцию, обобщив имеющиеся данные в области физиологии животных, профилактики заболеваний с определением оптимальных доз принудительного движения.
Рецензенты:
- доктор биологических наук, профессор А.И. Газизова
- доктор биологических наук, профессор Р. Ш. Тайгузин
ВВЕДЕНИЕ
Перевод животноводства на промышленную основу пагубно влияет на организм животного, так как сопровождается высокой плотностью животных на единицу площади, загазованностью помещений, повышенной влажностью, гиподинамией. Что приводит к ограничению подвижности животных и вызывает нарушения физиологических и обменных процессов в организме, появлению кожных заболеваний и некробактериоза (копытной гнили), поражению опорно-двигательной, дыхательной, пищеварительной систем и других органов. В связи с этим резко снижается продуктивность животных, падают сроки их эксплуатации, нарушается воспроизводительная функция (Х.Б. Баймишев, 1984, 1985). Одновременно, при правильной организации промышленного животноводства можно предотвратить эти негативные последствия.
Животноводческий комплекс промышленного типа – это, крупное специализированное производство, где на основе техники и научной организации труда осуществляются непрерывные технологические процессы, обеспечивающие ритмичный выпуск продукции. Условия содержания, кормления, цикличность, зоотехнических операций, использование генетического разнообразия пород в промышленных комплексах предопределяют интенсивное выращивание скота. Поэтому в современных животноводческих комплексах используют лучшие мировые генетические ресурсы пород скота, новые методы селекции, создание на основе скрещивания и гибридизации синтетических генотипов, которые должны обеспечить устойчивый прогресс и повышение мясной и молочной продуктивности. Разрабатываются новые, более совершенные системы нормирования кормления и оценки питательности кормов.
В условиях промышленных комплексов животные становятся восприимчивыми к стрессовым нагрузкам, резко проявляются некоторые их физиологические особенности: ухудшение качества мяса, внезапная гибель из-за сердечной недостаточности, возрастание потерь при транспортировании. Содержание животных в условиях гиподинамии привело к появлению у них ослабленной двигательной активности. Обособленность выращивания в комплексах вызывает у животных обостренное чувство агрессивности, которое проявляется во время смешивания групп и выражается в ухудшении качества кожного покрова. В зависимости от чувствительности к нагрузкам при транспортировании встречаются различные случаи, которые начинаются с утомления животного, затем переходят в необратимые симптомы миопатии нагрузки, или стрессового синдрома.
Особенность разведения скота современными методами (быстрый рост и образование мускулатуры при одновременном ограничении возможности движения) приводит к повышенному обмену веществ. При этом наблюдается диспропорция между массой сердца и тела, между объемом крови и массой животного. Одной из причин, ведущей к гибели животных при транспортировании, является дегенерация мышц — беломышечная болезнь, протекающая в относительно спокойных условиях незаметно (т.е. в период откорма). Для понимания всего происходящего специалисту необходимо знать биологические особенности экологической физиологии животных.
Что изучает экологическая физиология животных? Экофизиология животных – это, раздел физиологии, изучающий зависимость функций животных от условий жизни и деятельности в различных физико-географических зонах, в разные периоды года, суток, фазы лунного и приливного ритмов; раскрывает физиологические основы приспособлений (адаптации физиологической) к природным факторам. Экологическая физиология тесно связана с экологией, хронобиологией, возрастной и эволюционной физиологией, физиологией сельскохозяйственных животных, этологией. Использует для исследования все уровни физиологической интеграции: надорганизменный (популяционный), организменный, органный и системный, клеточный и субклеточный (молекулярный). Формирование адаптаций исследуют методами изоляции организма от отдельных факторов среды (искусственного выращивание), сравнения параметров физиологических реакций у близких в систематическом отношении видов с разной экологической специализацией, например арктических, тропических и пустынных, наземных и полуводных организмов с различной мышечной активностью. Экологическая физиология опирается на данные биофизики и биохимии.
Экологическая физиология — это, наука, изучающая основы практических мер по акклиматизации полезных видов животных, породному районированию с.-х. животных, акклиматизации человека в новых климатических районах в связи с их освоением. Экологическая физиология изучает физиологические характеристики, которые определяют направленность приспособительных реакций у разных животных, населяющих определённые регионы.
В Казахстане и России научные исследования по экологической физиологии были начаты с 30-х годов XX века. Основные из них можно перечислить: эколого-физиологические исследования терморегуляции (А. Д. Слоним, О. П. Щербакова), адаптации к жаркому и холодному климату (А. Ю. Юнусов, И. А. Израэль) и горным условиям (А. Г. Гинецинский, Н. Н. Сиротинин, М. М. Миррахимов), к среде с различным газовым составом (Е. М. Крепс, З. И. Барбашёва), механизмов зимней спячки животных (Н. И. Калабухов).
Экологический подход применен при изучении нервной системы (Д. А. Бирюков), генетики поведения (Д. К. Беляев, М. Е. Лобашев). С 50-х гг. исследования по экологической физиологии расширились в связи с освоением новых территорий (Арктика, субарктика, пустынные и горные районы); созданы стационары на высокогорных, пустынных и морских станциях, лаборатории при заповедниках. Актуальным становится изучение влияния шума, вибрации, температурных факторов на организм человека в целом и на функции отдельных органов (В. В. Парин, А. С. Трошин).
Различные проблемы экологической физиологии (environmental physiology) разрабатываются за рубежом: сравнительная физиология дыхания и мышечной деятельности (А. Крог), влияние пониженного атмосферного давления на дыхательные функции крови (Дж. Баркрофт), адаптация к низким температурам (К. Л. Проссер, Т. Х. Баллок, П. Ф. Шоландер; США), эколого-физиологические приспособления животных к жизни в пустыне (К. Шмидт-Нильсен; США), физиология труда человека в экстремальных условиях севера и пустыни (О. Г. Эдхолм; Великобритания). С 70-х гг. проблемы Экологическая физиология переплетаются с задачами охраны окружающей среды.
В последние десятилетия большое значение придаётся биологическим наукам, в том числе морфологии, изучающим строение организма животных в связи с гиподинамией и окружающей средой, которые накладывают отпечаток на строение и структуру тканей, органов, и системы организма. К числу таких факторов относятся загрязнения окружающей среды тяжелыми металлами, нефтепродуктами, газами, отходами атомной, тяжелой промышленности и другие (А.П. Жуков, 2000, 2001).
У животных опорно-двигательный аппарат сформировался под действием динамических нагрузок, особенно у копытных. При гиподинамии на него воздействуют, в основном статические нагрузки и уменьшаются динамические. По мнению ряда авторов (Х.Б. Баймишев, И.В. Хрусталёва, 1988; Б.П. Шевченко, Н.А. Малков, М.В. Андреев, 1992; Н.С. Кухаренко, 1992; Б.П. Шевченко, 1992; М.Ж. Нурушев, 2005) это является одной из основных причин появления остеомаляции, остеопороза, остеодистрофии, артритов, синовитов и миозитов. Повышенная влажность приводит к развитию некробактериоза.
Болезни копыт, от общего заболевания животных в промышленных комплексах, составляют 30%, а от общего количества животных в стаде и
сезонам года – до 70%. Учитывая, что на изменение функциональной нагрузки опорно-двигательного аппарата, первой при гиподинамии реагирует сердечнососудистая система, особенно вены – как наиболее её пластичное звено, а затем кости опорно-двигательной системы (Х.Б.Баймишев, 1985). Причем, анализ специальной литературы по данной теме не дал результатов.
Целью настоящего учебного пособия является, обобщить исследования российско-казахстанских ученых в решении проблем влияния различной двигательной активности и возраста на кровеносные сосуды и кости конечностей крупного рогатого скота, находящегося в условиях промышленного комплекса в онтогенезе, на:
1 – магистральные артерии организма крупного рогатого скота;
2 – магистральные вены организма и клапанный аппарат конечностей;
3 – экстраорганные вены фаланг пальцев и их венозные магистрали;
4 – влияние гиподинамии на клапанный аппарат вен, толщину стенок костных вен, в зависимости от места выхода их из костной ткани фаланг пальцев;
5 – характер опоры (площадь опоры, угол наклона) копытец;
6 – взаимосвязь вен копытной кости с венами мякиша и венозным руслом;
7 – разработать оптимальные дозы принудительного движения для молодняка крупного рогатого скота, в условиях промышленного комплекса.
Все указанные вопросы являются весьма актуальными и потому были подвержены нами детальному анализу на страницах данного научного труда. Необходимость обобщения результатов научных исследований по данной проблеме назрела давно. В ней ощущалась острая нужда и ученых и практиков. Солидарный труд ученых биологов двух стран позволил в определенной степени решить поставленную задачу. Безусловно, мы понимаем, что с разработкой и внедрением в науку новых методов исследования, появятся новые данные, которые расширят наше представление о гиподинамии.
Глубоко осознавая все плюсы многолетнего научного труда, посвященной изучению экологической физиологии животных по части кровеносных сосудов и периферического скелета крупного рогатого скота при гипо- и гипердинамии, мы надеемся, что эта книга, даст новый импульс в развитии отрасли молочного и мясного скотоводства. Считаем и верим, что она окажется полезной для студентов, аспирантов, магистрантов, докторантов, работникам АПК зооветеринарных специальностей и научным сотрудникам, занимающихся по данной проблеме.
По Государственному стандарту образования дисциплина «Экологическая физиология» предназначена для студентов специализации «экология» и «экологическая физиология человека и животных». Основной целью освоения дисциплины является формирование системных представлений о функционировании организма при различных воздействиях окружающей среды.
Для достижения поставленной цели выделяются задачи курса:
1) активировать знания базовых курсов физиологии и биохимии;
2) от уровня рассмотрения функций в среднестатистическом варианте перейти на уровень их изучения при реальных разнообразных средовых воздействиях;
3) рассмотреть основные представления об адаптациях и механизмы адаптаций к конкретным средовым факторам и их комплексам.
По окончании изучения указанной дисциплины студент должен иметь на современном уровне общее представление о:
- цели, задачах и методах изучения экологической физиологии, об основных подходах к исследованию адаптаций, о механизмах адаптации живых организмов к среде обитания, а также об особенностях адаптации к конкретным средовым факторам;
- знать основные факты, иллюстрирующие вышеперечисленные представления;
- уметь использовать полученные знания для обобщений и анализа новой информации.
1. СЕРДЕЧНОСОСУДИСТАЯ СИСТЕМА
Сердечнососудистая система и её придаток лимфатическая выполняют ряд функций в организме животных:
В целом кровеносная система представлена сердцем, артериями, венами и микроциркуляторным руслом (В.В.Куприянов, Я.Л. Караганов, В.И.Козлов, 1975;. Б.П.Шевченко, 1979, 1984, 1999; М.Ж. Нурушев, Б.П.Шевченко, М.С.Сеитов, А.Г.Гончаров, Х.Б. Баймишев, 2011). В функциональном отношении сердце обеспечивает движение крови по замкнутой системе кровеносных сосудов.
Вопросы для контроля
- Перечислите функции сердечной сосудистой системы?
- Какие системы относятся к интегративным?
- Что представляет кровь, лимфа?
- Как образуется внутренняя жидкостная среда организма?
- Где образуется и как разносится теплопродукция в организме?
- К чему приводит общее повышение температуры тела?
- Дайте морфологический состав сердечной сосудистой системы?
- Какие структуры встречаются в артериях и венах?
1.1. СЕРДЦЕ
Сердце в онтогенезе закладывается из двух соматических ангиобласт, преобразующиеся на правую и левую первичные грудные аорты. Оба сосуда, располагаясь снизу, справа и слева от хорды, вскоре сливаются в непарную грудную аорту, из которой впоследствии разовьётся сердце.
На стадии зародыша крупного рогатого скота шести сомитов сердце имеет трубчатую форму, 9 - 10 исчезает дорсальный мезокард, 10 – 12 сомитов сердце приобретает S-образную форму. На стадии зародыша 20 сомитов сердце крупного рогатого скота уже способно поддерживать циркуляцию крови по сосудам (Я.Т. Подковыров, 1967; В.К. Бирих, Г.М. Удовин, 1972). К концу предплодного периода развития оно превращается в четырёх камерное сердце.
Масса сердца плодов, после рождения молодняка и взрослых животных до 10 лет возрастает неравномерно и имеет постоянно тенденцию к увеличению. Абсолютный прирост массы сердца в плодном периоде развития значительно ниже, чем после рождения. Так в возрасте плодов 5 месяцев прирост массы сердца составил 23,06 г, а в 9 месяцев – 78,0 г. Однако эти цифры слабо характеризуют развитие сердца плодов. Относительные величины показывают наоборот, что за первую половину плодного развития масса сердца увеличивается в 116,3, а за вторую – только в 5,2 раза. Относительно к массе тела плодов сердце к рождению с 0,1% - в возрасте два месяца повышается до 0,94 % - девятимесячных плодов. О чём эти цифры свидетельствуют?
В.Р.Тарасов (1972) пишет, что разница между артериальным и венозным давлением плода незначительная, так как его сердце работает по типу присасывающего и нагнетающего насоса. Недостаток кислорода в крови плода компенсируется сокращением сердца в 2 – 3 раза чаще, чем у взрослых животных. Из этого следует, что с возрастом плодов и формированием органов, систем и началом их функционирования нагрузка на сердце повышается.
Повышение нагрузки ведёт к росту массы сердца, а рост массы приводит к повышению его функции. Эти два понятия не разделимы, они постоянно влияют друг на друга и взаимосвязаны (табл. 1).
1. Рост массы сердца крупного рогатого скота
|
Возраст, мес. |
Живая масса тела, кг. |
Масса сердца, г. |
Абсолют. прирост, г. |
Относит. рост, раз. |
В % к массе тела |
|
Плоды 2 |
0,02 |
0,2 |
- |
- |
0,1 |
|
3 |
0,18 |
0,45 |
0,25 |
2,25 |
0,25 |
|
4 |
1,01 |
7,76 |
7,31 |
17,24 |
0,70 |
|
5 |
3,22 |
23,26 |
15,50 |
3,00 |
0,72 |
|
2 - 5 |
- |
- |
23,06 |
16,70 |
- |
|
6 |
5, 15 |
40,40 |
17,14 |
1,73 |
0,78 |
|
7 |
8,20 |
64,00 |
23,60 |
1,60 |
0,79 |
|
8 |
15,25 |
130,70 |
66,70 |
2,04 |
0,85 |
|
9 |
22,10 |
208,70 |
78,00 |
1,60 |
0,94 |
|
6 - 9 |
- |
- |
168,30 |
5,20 |
- |
|
Новорож. |
30,40 |
212,00 |
3,30 |
1,02 |
0,70 |
|
1 мес. |
36,90 |
283,10 |
71,10 |
1,34 |
0,76 |
|
2 |
46,60 |
330,60 |
47,50 |
1,17 |
0,71 |
|
3 |
65,60 |
380,30 |
49,70 |
1,15 |
0,58 |
|
6 |
145,90 |
715,80 |
335,50 |
1,88 |
0,49 |
|
1 - 6 |
- |
- |
503,80 |
3,38 |
- |
|
12 |
215,00 |
980,00 |
264,20 |
1,37 |
0,46 |
|
18 |
263,00 |
1218,00 |
238,00 |
1,24 |
0,46 |
|
6 - 18 |
- |
- |
502,20 |
1,70 |
- |
|
24 |
300,00 |
1360,00 |
141,20 |
1,12 |
0,45 |
|
36 |
327,30 |
1439,00 |
79,00 |
1,11 |
0,44 |
|
48 |
376,00 |
1816,70 |
377,70 |
1,26 |
0,48 |
|
60 |
408,50 |
1949,00 |
132,30 |
1,07 |
0,48 |
|
120 |
426,70 |
2057,70 |
108,70 |
1,05 |
0,48 |
После рождения абсолютный прирост массы сердца молодняка постепенно возрастает и в шесть месяцев составляет 503,8 г, в среднем в месяц увеличивается на 83, 9 г, напротив, с 6 до 18 месяцев (за 12 месяцев) только 502,2 г, т е. в месяц – 41,8 г, что в два раза ниже, чем за первое полугодие жизни молодняка. Но относительно к массе тела сердце, как и плодов, сохраняется до возраста телят двух месяцев (0,71 %). Затем плавно понижается до 6 месяцев и далее, независимо от роста массы тела взрослых животных составляет 0,46 – 0,48 %, то есть относительно к массе тела масса сердца поздних плодов и телят после рождения до трёх месяцев выше (табл.1).
И так сердце является центральным органом кровеносной системы, которое нагнетает кровь в артериальную систему животных и обеспечивает совместно с мышцами обратный её возврат к сердцу по венам.
Вопросы для контроля
- Из чего и как формируется аорта?
- Как первичные две аорты преобразуются в сердце?
- На какой стадии развития зародыша сердце имеет S-образную форму?
- Какую функцию выполняет сердце?
- Как изменяется масса плодов и животных после рождения?
- Какой пульс плода и животного после рождения?
- Как влияет рост массы сердца на его функцию или наоборот?
1.2. КРОВЕНОСНЫЕ СОСУДЫ
Кровеносные сосуды закладываются из соматических и желточных ангиобласт. Впоследствии они преобразуются в кровеносные сосуды, представляющие систему сообщающихся трубок и подразделяются, на артерии, вены и лежащее между ними микроциркуляторное русло. Артерии несут кровь от сердца, вены – к сердцу, независимо какая по ним течёт кровь (табл.2).
Стенка артерий состоит интимы, медии и экстерны. Интима представлена эндотелиоцитами, подэпителиальной (базальной) мембраной. Рельеф интимы изнутри покрыт гликокаликсом (Ю.В.Котелевцев, 2011) и представлен первичными, вторичными микроскладками, которые являются резервом в случае гипертензии. Они могут играть значительную роль в гемодинамике крови.
Медия состоит из нескольких слоёв или отдельных мышечных клеток, лежащих чаще под косым или прямым углом по отношению к продольной оси сосуда, между пучками которых располагаются ретикулярные волокна, окончиваяющиеся в эластической мембране. В итоге формируется строма кровеносного сосуда. Экстерна – это рыхлая соединительная ткань, в которой проходят отдельные волокна и пучки симпатической части вегетативной нервной системы и vasa - vasorum. В стенке сосудов часто встречаются баро-, термо- и хеморецепторы и т.д.(рис. 1).
2. Отличительные признаки кровеносных сосудов
|
№ п/п |
Артерии |
Вены |
|
1. |
На поперечном срезе зияют |
На поперечном срезе спадаются |
|
2. |
Стенка толстая |
Стенка тонкая |
|
3. |
Из артерий на срезе живых животных фонтанирует алая кровь |
Из вен изливается кровь тёмно-вишнёвого цвета |
|
4. |
В артериях нет клапанов, исключение составляет луковица аорты |
В венах имеются одно-, двух- и трёхстворчатые клапаны, встречаются трабекулы, пластинки, складки, лакуны, мысы и др. |
|
5. |
В стенках артерий нет жомов, |
Имеются жомы (сфинктеры) |
|
6. |
В сосудисто-нервных пучках артерия одна |
Вен может быть две и даже более |
|
7. |
Артерии меньше по диаметру вен |
Диаметр вен взрослых животных в 1,8 – 2,2 раза больше диаметра артерий |
|
8. |
Артерии образуют сети |
Вены сети, сплетения, кавернозные Тела |
Примечание: Есть и другие отличительные признаки, но эти наиболее частые.

Рис.1. Стенка артерии с широким просветом. Импрегнация азотнокислым серебром. Об. 90, ок. 7.
По толщине и количеству эластических, сократительных элементов в стенке, артерии подразделяются: на артерии эластического, смешанного и мышечного типа и несут кровь богатую питательными веществами, кислородом. К артериям эластического типа относится луковица аорты, к смешанному – дуга, грудная и брюшная аорта, мышечного типа – подключичная, подмышечная, общая сонная, наружная подвздошная, бедренная артерии и другие (рис.2).
Рис.2. Артерия мышечного типа с узким просветом.
Импрегнация азотнокислым серебром. Об.90, ок.7.
Вены, как правило, несут кровь богатую двуокисью углерода, продуктами метаболизма в правое предсердие, органы нейтрализации и выделения Исключение составляют легочные вены, по ним течёт артериальная кровь из лёгких в левое предсердие. Вены - наиболее пластичное звено кровеносной системы, чаще подвергающиеся изменениям под действием различных экологических, динамических, химических и других факторов.
Стенка вен тоньше, чем артерий, и подразделяется на интиму, медию и экстерну, но наружная соединительнотканная оболочка толще, чем интима и медия. Вены конечностей, головы и шеи жвачных более богато снабжены мышечными элементами, чем вены тела, располагающиеся в горизонтальной плоскости.
К главным воротам мышц, как правило, направляется триада: артерия, вена и нерв. Чаще вена располагается между артерией и нервом. В серозных оболочках, фасциях артерия может находиться между двумя венами, что способствует оттоку крови по ним за счёт пробегающей систолической волны по артериям (Б.П.Шевченко, 1975, 1976). В костных каналах (подглазничном, нижнечелюстном, подошвенном - третьей фаланги пальцев и др.) диаметр артерий равен калибру вен и вены в них спиралевидно оплетают артерии, что способствует движению крови в них под действием систолической волны сердца. В просвете вен часто встречаются внутривенные образования: клапаны, трабекулы, пластинки, складки, лакуны, мысы, играющие важную роль в гемодинамике. Иннервирует сосуды симпатическая часть вегетативной нервной системы.
Вопросы для контроля
- Из чего закладываются кровеносные сосуды?
- Укажите деление кровеносных сосудов?
- Какими структурами представлена стенка сосудов?
- Чем представлена внутренняя оболочки стенки сосудов?
- Чем представлена средняя оболочка стенки сосудов?
- Чем образована наружная оболочка сосудов, какие структуры?
- Какие нервные окончания встречаются в стенке сосудов?
- Какие отличительные признаки артерий от вен?
- На какие типы делятся артерии?
- Какая кровь течёт по венам?
- По каким венам течёт артериальная кровь?
- Что такое «триада»?
- Какие структуры встречаются в просвете вен?
1.3. МИКРОЦИРКУЛЯТОРНОЕ РУСЛО
Микроциркуляторное русло объединяет артерии и вены в систему сообщающихся трубок и рассматривается как специализированный эффектор, обеспечивающий трофическую, регуляторную, интегративную и защитную функции (О.В.Алексеев, 1981). Через него происходит фильтрация жидкости с питательными веществами и химическими элементами из крови в ткани, и обратно всасываются продукты метаболизма, отработанные соли и т.д. В итоге, через стенки сосудов микроциркуляторного русла в ткани растущего организма проникают органозоли, формирующие жидкую химическую среду организма - гомеостаз и «поле роста» для развития, размножения клеток, тканей, органов, которые в целом обеспечивают рост организма (Б.П.Шевченко 1975). Воздействие этой среды, особенно на развивающийся организм громадное. Она может при силосном, сенажном кормлении, в результате накопления токсических веществ в организме, привести к подавлению развития плода, или патологическому искажению и даже к смерти (Б.П.Токин, 1970; П.Г.Светлов, 1978; Б.П.Шевченко, 1984, 1999).
Микроциркуляторное русло подразделяется на звено притока (артерии), распределения (артериолы, прекапилляры и их сфинктеры), обмена (капилляры) и дренажное депонирующее звено (посткапилляры, венулы, рис.3).
Рис. 3. Микроциркуляторное русло фасции. Импрегнация
азотнокислым серебром. Об.20, ок. 10.
Длина капилляров может достигать 1,5 – 3,0 мм, скорость тока крови в них в пределах 0,5 мм/сек. Площадь капилляров больше площади поперечного сечения аорты в 800 – 1000 раз. По этому, скорость тока крови в них должна быть ниже, чем в аорте приблизительно в 1000 раз. Все звенья, составляющие микроциркуляторное русло, имеют свои особенности строения и функции (В.В.Куприянов, Я.Л.Караганов, В.И.Козлов, 1975, Б.П.Шевченко, 1979).
Обсуждение звеньев микроциркуляторного русла было бы неполным без анализа артериовенозных анастомозов (АВА). В настоящее время АВА за редким исключением (суставной хрящ, роговица глаза, эпителий), обнаружены почти во всех органах. Через них, в случае покоя органа, происходит сброс
крови из артерий в вены, минуя микроциркуляторное русло. Они бывают гломусного, замыкательного (шунты) и магистрального типов строения (Ю.
Т. Техвер, 1972, 1989).
АВА гломусного типа представляют извилистый конгломерат сосудов, стенка которых не имеет эластической мембраны и представлена интимой, медией и экстерной, большим числом хемо-, баро-, терморецепторов. Они могут открываться в одну или несколько вен.
АВА замыкательного типа чаще бывают шунтами и полушунтами. Шунты прямо соединяют артерию с веной и, через них, кровь непосредственно сбрасывается в вены, по этой причине они короткие и характеризуются наличием хорошо развитой меди. В соустье с артериолами и венулами мышечные клетки ориентированы циркулярно, в промежуточном сегменте – косо, под острым углом относительно продольной оси шунта. Подобное расположение мышечных клеток указывает на различную их функцию (рис.4).
Полушунты по форме могут быть дуговыми, прямыми и по длине превосходят шунты. Артериальный сегмент их заужен, а венозный – расширен и без миоцитов. Такие АВА могут отдавать прекапилляры, капилляры, образующие обменные сети.

Рис.4. Артериоловенулярный анастомоз типа шунта с двумя звеньями. Импрегнация азотнокислым серебром. Об. 20, ок.12,5.
АВА с помощью сократительных элементов регулирует кровоток в русле, направляя его либо в капиллярные сети во время активного функционирования органов, либо, минуя, сбрасывают кровь непосредственно в вены, во время покойного состояния мышц. В связи с наличием в стенках АВА различных рецепторных окончаний, они очень чувствительны к температурным, химическим и механическим воздействиям. По этой причине, АВА чаще встречаются в коже, стенках пищеварительной, дыхательной систем, в миокарде,
в серозных оболочках и других органах. В настоящее время они обнаружены в почти во всех органах.
1.4. К ВОПРОСУ ТИПОВОЙ АНАТОМИИ КРОВЕНОСНЫХ СОСУДОВ
Тип ветвления кровеносных сосудов имеет большое практическое значение
при выборе доступа оперативного вмешательства на органе, наложении лигатур, решения вопросов гемодинамики и так далее. При рассыпном типе ветвления сосудов - условия возникновения коллатерального кровообращения хуже, чем при магистральном (Б.А.Долго-Сабуров, 1961). Методами препарирования плечеголовного ствола животных установлены три типа его ветвления: ассиметричный, симметричный и переходный. Чаще встречается симметричный тип ветвления, реже оба крайних. Для крупного рогатого скота тип ветвления дуги аорты не установлен.
Известны три типа ветвления сосудов: первый тип – рассыпной, второй – магистральный и третий – переходный. Согласуя учение П.Ф.Лесгафта (1909) о соответствии деления основных стволов артерий с костной основой тела животных, А.И. Акаевский (1975 и последующие издания) привёл деление артерий в соответствии с костными сегментами и назвал их ангиомерами. К двум крайним типам ветвления В.Н. Шевкуненко, А.И. Геселевич предложили ещё два: дихотомический и концевой (магистральный), потом появится трихотомический.
По типам ветвления кровеносных сосудов в научной литературе нет ещё единого мнения. Сущность классификации, предложенной С.Н.Касаткиным (1969) заключается в том, что по артериальному стволу различают унитрункальные, бифуркационные, трифуркационные и мультифидные, вены – унизоматные (моноконфлюэнтные), биконфлюэнтные, триконфлюэнтные и поли(мульти)конфлюэнтные. Типы артерий и вен соединяются со звеньями микроциркуляторного русла определённого диаметра и частоты. Поэтому это не праздный вопрос. В настоящее время приняты типы ветвления для артерий: магистральный, дихотомический, трихотомический и рассыпной; для вен – моно-, би-, три- и поликонфлюэнтный типы.
1.5. ОКОЛЬНОЕ КРОВООБРАЩЕНИЕ В СИСТЕМЕ СОСУДОВ
Более углублённое исследование различных анастомозов кровеносных сосудов человека проводил Б.А.Долго-Сабуров (1961), которое вылилось в учение о коллатеральном кровообращении. Позднее было установлено, что кровеносная система обладает колоссальными резервными возможностями,
большой пластичностью и приспособляемостью к изменившимся условиям кровообращения под действием различных внешних и внутренних факторов (Х.Б.Баймишев, 1983, 1984, 1985, 1987; И.В.Хрусталёва, Б.В.Криштофорова, Л.Г.Харченко, Д.А.Черепахина, 1987, Б.П.Шевченко, 1999 и др.).
Артерии и вены могут соединяться между собой по типу «конец в конец», «конец в бок», «бок в бок» и т.д. В итоге артерии чаще образуют сети, а вены - сети, сплетения, кавернозные тела, но в формировании последних могут принимать участие и артерии (Б.П.Шевченко, 1999).
Экспериментальным путём было установлено, что при наложении лигатур на артерии образуются через ветви отходящие от культи, различные анастомозы, коллатерали. Было сделано заключение, что прямые анастомозы развиваются не только из капилляров культи магистральных артерий, но источником их образования могут служить капилляры рядом расположенных органов, чаще фасций, врастающие в область культи (Б.А. Долго-Сабуров, 1961).
В системе сосудов бывают внутрисистемные и межсистемные анастомозы. Они часто встречаются в организме животных в области шеи и головы, грудной и брюшной полостей, на конечностях, особенно, в области суставов. В артериальном круге головного мозга, в стенках его сосудов заложены контрактильные приспособления (жомы) с рецепторными и двигательными окончаниями, которые регулируют кровоток как в системе сосудов правой, так и левой стороны головного мозга (И.В.Ганнушкина, 1973).
В целом необходимо помнить, что окольное кровообращение в системе сосудов животных изучено крайне слабо, а у некоторых по разным причинам вообще не исследовалось.
Вопросы для контроля
- Что представляет и какую функцию выполняет микроциркуляторное русло?
- Что такое гомеостаз?
- На какие структуры делится микроциркуляторное русло?
- Что такое АВА и какого типа они бывают?
- Особенности строения АВА?
- Какие типы ветвления артерий и вен приняты в настоящее время?
- Что такое окольное кровообращение?
- Что такое анастомозы?
- Какие бывают анастомозы?
- Что такое «жомы» и где они чаще встречаются?
2. АОРТА
Аорта основной сосуд большого круга кровообращения, выходящий из аортального отверстия левого желудочка сердца. В аорте кровь движется со
скоростью 50 см/сек, а в капиллярах – 0,5 мм/сек. Топографически она делится у животных на дугу аорты, грудную и брюшную аорту.
2.1. ОСОБЕННОСТИ РОСТА ДУГИ АОРТЫ
Дугу аорты плодов и животных топографически и функционально можно разделить на три сегмента:
1 – от аортального отверстия сердца до отхождения плечеголовного ствола, в который входит луковица аорты;
2 - от плечеголовного ствола до отверстия артериального протока плода или артериальной связки у взрослых животных;
3 – от артериального протока (связки) до пятого – шестого грудного позвонка.
В различные возрастные периоды, каждый сегмент характеризуется различной интенсивностью роста диаметра (Я.Т.Подковыров, 1969; В.К.Бирих, Г.М.Удовин, 1972, Б.П.Шевченко, 1987). Кроме этого на их калибр роста в плодном периоде развития влияет диаметр артериального протока (табл.3).
3. Рост диаметра (мм) сегментов дуги аорты
|
Возраст. мес. |
С е г м е н т ы |
Артериальный проток (связка) |
|||||||
|
1 |
2 |
3 |
|||||||
|
Плодный период развития |
|||||||||
|
3 |
3,20 + 0,21 |
3,25 + 0,30 |
3,64 + 0,18 |
4,70 + 0,18 |
|||||
|
4 |
4,36 + 0,14 |
3,90 + 0,11 |
5,40 + 0,10 |
5,70 + 0,17 |
|||||
|
5 |
8,63 + 0,22 |
7,05 + 0,36 |
7,83 + 0,27 |
7,82 + 0,29 |
|||||
|
6 |
11,24 + 0,32 |
9,20 + 0,32 |
10,10 + 0,22 |
9,45 + 0,25 |
|||||
|
7 |
14,14 + 0,38 |
10,52 + 0,42 |
13,40 + 0,38 |
10,65 + 0,18 |
|||||
|
8 |
17,36 + 0,21 |
12,77 + 0,36 |
16,36 + 0,21 |
13,75 + 0,35 |
|||||
|
9 |
20,22 + 0,24 |
16,60 + 0,55 |
18,10 + 0,24 |
12,85 + 0,52 |
|||||
|
Постнатальный период развития |
|||||||||
|
3 – 7 дн. |
21,50 + 0,36 |
19,30 + 0,23 |
20,50 + 0,36 |
9,95 + 0,59 |
|||||
|
1 мес. |
24,40 + 0,29 |
23,00 + 0,35 |
23,40 + 0,29 |
5,34 + 0,65 |
|||||
|
6 |
26,00 + 0,24 |
26,20 + 0,18 |
26,00 + 0,20 |
Связка |
|||||
|
12 |
28,65 + 0,35 |
28,40 + 0,22 |
28,55 + 0,25 |
|
|||||
|
18 |
32,17 + 0,28 |
31,95 + 0,24 |
31,17 + 0,24 |
|
|||||
Диаметр первого сегмента замерялся на уровне верхних краёв створок клапанов аорты, второго – в центре между плечеголовным стволом и отверстием артериального протока (связки), третьего – на уровне V-VI грудного позвонка (в точке отхождения первой спинномозговой артерии). Калибр артериального протока снимался в центре, так как его соустья расширены, а центр - суженный. Сосуды наполнялись латексом и массой АКР.
В различные возрастные периоды диаметр каждого из этих сегментов характеризуется различной активностью роста. Калибр первого и третьего сегмента возрастает более равномерно относительно друг друга, а второго – до возраста одного месяца после рождения отстаёт в развитии по отношению к первому и третьему сегментам. Диаметр артериального протока до возраста плодов восьми месяцев увеличивается интенсивнее второго сегмента, о чём свидетельствуют данные таблицы 3, но, начиная с восьми месяцев плодного развития и до месячного возраста после рождения, он постепенно зауживается и у телят в возрасте 1,5 - 2 месяца превращается в связку. К возрасту молодняка шесть месяцев диаметр второго сегмента выравнивается с первым и третьим.
4. Абсолютный прирост диаметра (мм) сегментов дуги аорты
|
Возраст. мес. |
С е г м е н т ы |
Артериальный проток (связка) |
|||||||
|
1 |
2 |
3 |
|||||||
|
Плодный период развития |
|||||||||
|
4 |
1,16 |
0,67 |
1,76 |
1,0 |
|||||
|
5 |
4,27 |
3,15 |
2,43 |
2,12 |
|||||
|
6 |
2,61 |
2,15 |
2,27 |
1,73 |
|||||
|
3-6 |
7,34 |
5,95 |
6,46 |
4,85 |
|||||
|
7 |
2,90 |
1,32 |
3,30 |
1,10 |
|||||
|
8 |
3,22 |
2,25 |
2,96 |
3,10 |
|||||
|
9 |
2,86 |
3,83 |
1,74 |
0,90 |
|||||
|
6-9 |
8,98 |
7,40 |
8,00 |
3,30 |
|||||
|
Постнатальный период развития |
|||||||||
|
3 – 7 дн. |
1,28 |
2,70 |
2,40 |
-2,90 |
|||||
|
1 мес. |
2,90 |
3,70 |
2,90 |
-4,61 |
|||||
|
6 |
1,60 |
3,20 |
2,60 |
- |
|||||
|
3 дн. – 6 мес. |
4,50 |
6,90 |
5,50 |
- |
|||||
|
12 |
2,65 |
2,20 |
2,55 |
- |
|||||
|
18 |
3,52 |
3,55 |
2,62 |
- |
|||||
Абсолютный прирост диаметра сегментов с возрастом плодов и животных после рождения увеличивается неодинаково. В первую половину плодного развития (3–6 месяцев) наиболее активно возрастает диаметр первого и третьего сегментов, а второго - отстаёт. Эта закономерность в приросте диаметров сохраняется и до рождения, но диаметр артериального протока активнее увеличивался в 3 – 4 и 8 месяцев плодного развития и слабее до девяти (табл.4).
После рождения до возраста молодняка шести месяцев очень активно увеличивается калибр второго сегмента. С 12 месяцев и старше прирост диаметра всех сегментов аорты был равномерным.
Проблема морфогенеза сердечнососудистой системы включает большое число различных вопросов: от образования ангиоостровков (ангиобласт) и до преобразования их с возрастом в органы кровеносной системы. Нет ни одной стадии онтогенеза, на протяжении которой, были бы исключены динамические изменения формы и структуры. Поэтому преобразования по стадиям онтогенеза, имеют не только теоретическое, но и важное практическое значение, знание которых поможет избежать воздействия неблагоприятных факторов на развивающийся организм - как в плодном, так и в постнатальном периодах развития
В животном организме сердечнососудистая система функционирует по законам замкнутой системы трубок. Артерии выполняют не только механическую функцию проведения крови до микроциркуляторного русла, но они активно воздействуют на гемодинамику и давление в них. Вены не только возвращают кровь с периферии организма к сердцу, но и способны накапливать её. Таким образом через накопление крови в системе вен, она может вызвать гипертензию (В. Folkov, W. Niel, 1976).
Описанные сегменты с возрастом животных, увеличивают диаметр неравномерно как в целом, составляющие дугу, так и отдельно каждый сегмент. Особенно большим колебаниям подвержен второй сегмент относительно первого и третьего, и артериального протока. Из приведённого анализа следует, что артериальный проток относительно третьего сегмента дуги аорты у плодов выполняет большую функциональную нагрузку, чем второй сегмент, наименьшие размеры диаметра, которого в плодном периоде развития способствуют местному сопротивлению тока крови от левого желудочка сердца в грудную аорту. Сформированная таким образом «горловина», располагающаяся на месте второго сегмента дуги, содействует интенсивному кровотоку из первого сегмента дуги аорты в плечеголовной ствол. Одновременно поток крови, оттекающий из артериального протока в грудную аорту, способствует току крови из первого сегмента в третий через «горловину» по принципу инжектора. Отсюда следует, что второй сегмент выполняет с одной стороны функцию анастомоза, а с другой – биологического регулятора, направляющего кровь через плечеголовной ствол в краниальную часть тела для развития, прежде всего, головного мозга и передней части тела плода.
Из сравнения диаметра второго сегмента дуги аорты и артериального протока видно, что диаметр их относительно друг друга увеличивается волнообразно. С 8-месячного возраста плодов диаметр артериального протока суживается и к возрасту телят 1,5 – 2 месяца запустевает (облитерирует), превращаясь в связку.
Таким образом, дуга аорты имеет не только сложное происхождение, но и сложный онтогенез адаптивных сегментов.
Вопросы для контроля
- На какие сегменты делится дуга аорты животных?
- Объясните рост сегментов дуги аорты?
- Какую систему представляют кровеносные сосуды?
- Что такое артериальный проток и когда он функционирует?
- Какие части тела снабжают кровью плечевой ствол?
- По каким законам функционирует кровеносная система?
2.2. РОСТ ДИАМЕТРА ПЛЕЧЕГОЛОВНОГО СТВОЛА
Плечеголовной ствол снабжает кровью органы шеи, головы, грудных стенок и конечностей. Что бы можно было полно судить о его морфофункциональных изменениях, мы провели измерения всего комплекса его сосудов (рис.5).
Наружный диаметр плечеголовного ствола плодов крупного рогатого скота с возрастом увеличивается равномерно, по сравнению с сегментами дуги аорты. Некоторая активизация роста пиков его размеров отмечается в возрасте плода 5 и 7 месяцев. После рождения его калибр активно возрастает у новорожденных и телят в шесть месяцев. Равномерный рост диаметра ствола продолжается до 18 месяцев, но темп роста его характеризуется более низкими показателями, чем у телят до 6 месяцев (табл. 5).

Рис.5. Ветвление плечеголовного ствола. Вид со стороны грудной полости.
1. плечеголовной ствол;
2. рёберно-шейный ствол;
3. левая и правая подключичные артерии;
4. внутренняя грудная;
5. поверхностная шейная;
6. наружная грудная артерии;
7. ствол сонных артерий;
8. общие сонные артерии;
9. подмышечная;
10. передняя межреберная;
11. дорсальная лопаточная;
12. глубокая шейная;
13. позвоночная;
14. нисходящая ветвь внутренней грудной артерии.
5. Рост диаметра (мм) плечеголовного ствола
|
Возраст мес. |
Диаметр внешний М ± m |
Абсолют. прирост |
Диаметр внутрен. М ± m |
Абсолют. прирост |
||
|
Плодный период развития |
||||||
|
3 |
2,10 + 0,12 |
- |
1,20 + 0,12 |
- |
||
|
4 |
3,10 + 0,12 |
1,00 |
1,60 + 0,14 |
0,60 |
||
|
5 |
5,30 + 0,26 |
2,20 |
4,40 + 0,16 |
2,80 |
||
|
6 |
6,90 + 0,28 |
1,60 |
5,00 + 0,09 |
0,60 |
||
|
3 – 6 |
- |
4,80 |
- |
3,80 |
||
|
7 |
9,00 + 0,26 |
2,10 |
8,90 + 0,40 |
1,90 |
||
|
8 |
10,70 + 0,27 |
1,70 |
7,90 + 0,11 |
1,00 |
||
|
9 |
11,90 + 0,07 |
1,20 |
9,70 + 0,13 |
1,80 |
||
|
6 – 9 |
- |
5,00 |
- |
4,70 |
||
|
Постнатальный период развития |
||||||
|
3 – 7 дн. |
13,10 + 0,20 |
1,20 |
10,70 + 0,14 |
1,00 |
||
|
1 мес. |
14,70 + 0,28 |
1,60 |
12,97 + 0,17 |
2,27 |
||
|
3 |
16,90 + 0,24 |
2,20 |
14,30 + 0,25 |
1,33 |
||
|
6 |
23,10 + 0,24 |
6,20 |
17,90 + 0,53 |
3,60 |
||
|
3 дн.- 6 мес. |
- |
10,00 |
- |
7,20. |
||
|
12 |
24,50 + 0,49 |
1,40 |
20,60 + 0,25 |
2,70 |
||
|
18 |
27,00 + 0,52 |
2,90 |
22,80 + 0,25 |
2,70 |
||
С возрастом плодов и телят после рождения внутренний просвет ствола изменяется несколько иначе. Так, в плодном периоде развития он наиболее активно увеличивается в 5, 7 и девять месяцев. После рождения темп роста его замедляется и соответствует развитию внешнего диаметра.
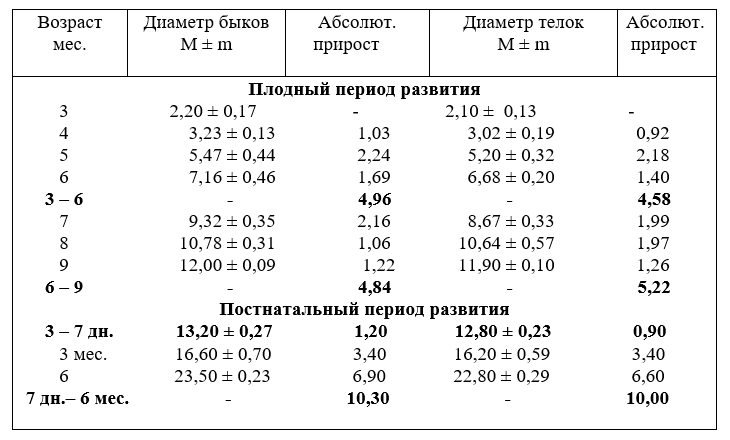
У разнополых животных диаметр плечеголовного ствола развивается неодинаково. Он активнее возрастает у быков и слабее – у тёлок. К возрасту одного года диаметр ствола быков и тёлок выравнивается и в старших возрастных группах остаётся стабильными (табл. 6).
Толщина стенок плечеголовного ствола толще в плодном периоде развития и тоньше – после рождения. Наиболее толстая стенка ствола у плодов в возрасте 3 и 4 месяца и составляет 45–48% от общего диаметра, в пять, шесть месяцев –27%, в девять – 18%. После рождения толщина стенок ствола значительно уступает толщине стенок в плодном периоде развития и достигает 16,8% от диаметра ствола.
6. Рост диаметра (мм) плечеголовного ствола разнополых животных
2.3. РОСТ ДИАМЕТРА ПОДКЛЮЧИЧНЫХ АРТЕРИЙ
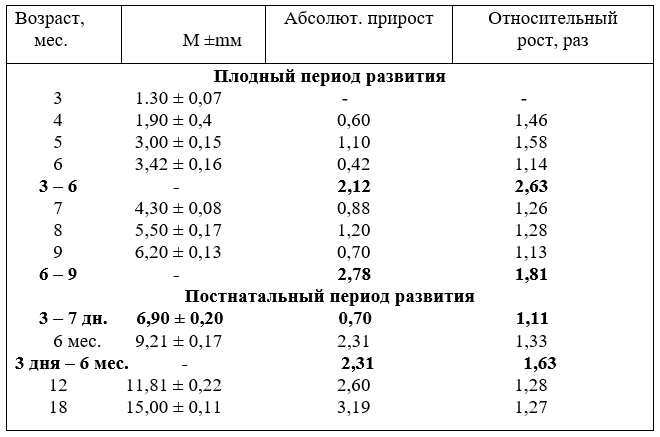
Диаметр подключичных артерий с возрастом плодов тоже увеличивается неравномерно. Наиболее активно он возрастает у плодов в пять. семь и восемь месяцев. За первую половину плодного развития абсолютный прирост диаметра составил 2,12 мм, а за вторую – 2,78 мм. В относительных величинах, наоборот, в первую половину плодного развития он увеличился в 2,63, а за вторую – в 1,81 раза (табл.7).
После рождения абсолютный прирост диаметра артерии активно возрастает до года и устанавливается в 18 месяцев. По отношению к плечеголовному стволу диаметр подключичной артерии изменяется несколько иначе. Так, у плодов трёх месяцев просвет подключичной артерии составляет 62,2% от диаметра плечеголовного ствола. К рождению он понижается до 51,9%. В связи с повышением функции грудных конечностей после рождения диаметр подключичной артерии относительно плечеголовного ствола у новорожденных увеличивается до 52,7%. а в годичном возрасте составляет 54,7%. Из этого следует, что самыми относительно широкими артерии остаются в возрасте плодов трёх месяцев.
7. Рост диаметра (мм) подключичных артерий
2.4. РОСТ ДИАМЕТРА СТВОЛА СОННЫХ АРТЕРИЙ
Ствол сонных артерий является непосредственным продолжением к голове плечеголовного ствола. Его диаметр превышает диаметр подключичных артерий в плодном периоде развития в среднем в 1,49, а в постнатальном – в 1,34 раза. Ствол сонных артерий встречался в 81,4 % случаев, а в 18,6% отсутствовал. В случае отсутствия от правой подключичной артерии последовательно отходили левая, а затем правая общие сонные артерии.
Диаметр ствола сонных артерий с возрастом плодов и животных увеличивается неравномерно. Наиболее активно он возрастает у плодов в возрасте пять, шесть, семь и девять месяцев. Особенно бурно диаметр ствола увеличивается у телят впервые дни постнатальной жизни. Затем темп роста замедляется, но до 18 месяцев всё-таки остаётся высоким и стабилизируется в этом возрасте, в последующие возрастные группы он увеличивается, но незначительно (табл. 8).
8. Рост диаметра (мм) ствола сонных артерий
С возрастом диаметр ствола сонных артерий относительно калибра плечеголовного ствола у плодов составляет, соответственно – 95,7% и у телят - 90,0%. То есть к рождению диаметр ствола суживается и неодинаково. Самый широкий просвет ствола сонных артерий наблюдается у плодов в возрасте три и четыре месяца и достигает в девять месяцев 73,7%.
После рождения относительно широкий просвет ствола остаётся у телят в возрасте до одного месяца, в три месяца составляет 73,7%, т.е. остаётся на уровне новорожденных телят. Затем до 18 месяцев наступает период волнообразного роста и снижения диаметра относительно плечеголовного ствола.
Из сравнения роста диаметра ствола сонных и подключичных артерий видно, что калибр ствола превосходит диаметр подключичных артерий у новорожденных в 1,46, у молодняка в шесть месяцев – в 1,41 и в 18 месяцев - в 1,1 раза.
Вопросы для контроля
- На какие артерии делится плечеголовной ствол?
- Как изменяется диаметр плечеголовного ствола?
- Как увеличивается диаметр плечеголовного ствола быков и телок?
- Какой сосуд следует считать продолжением к голове плечеголовного ствола?
2.5. РОСТ ДИАМЕТРА ОБЩИХ СОННЫХ АРТЕРИЙ
Общие сонные артерии располагаются в пищеводно-трахеальном жёлобе, во влагалище, сформированном листками глубокой фасции. Артерии плодов и молодняка снаружи прикрывает вилочковая железа, грудино-челюстная, грудино-сосцевидная, плечеголовная, подкожная мышца шеи и платизма. К медиодорсальной стенке артерий в фасциальном влагалище прилежит вагосимпатический ствол (рис.6).
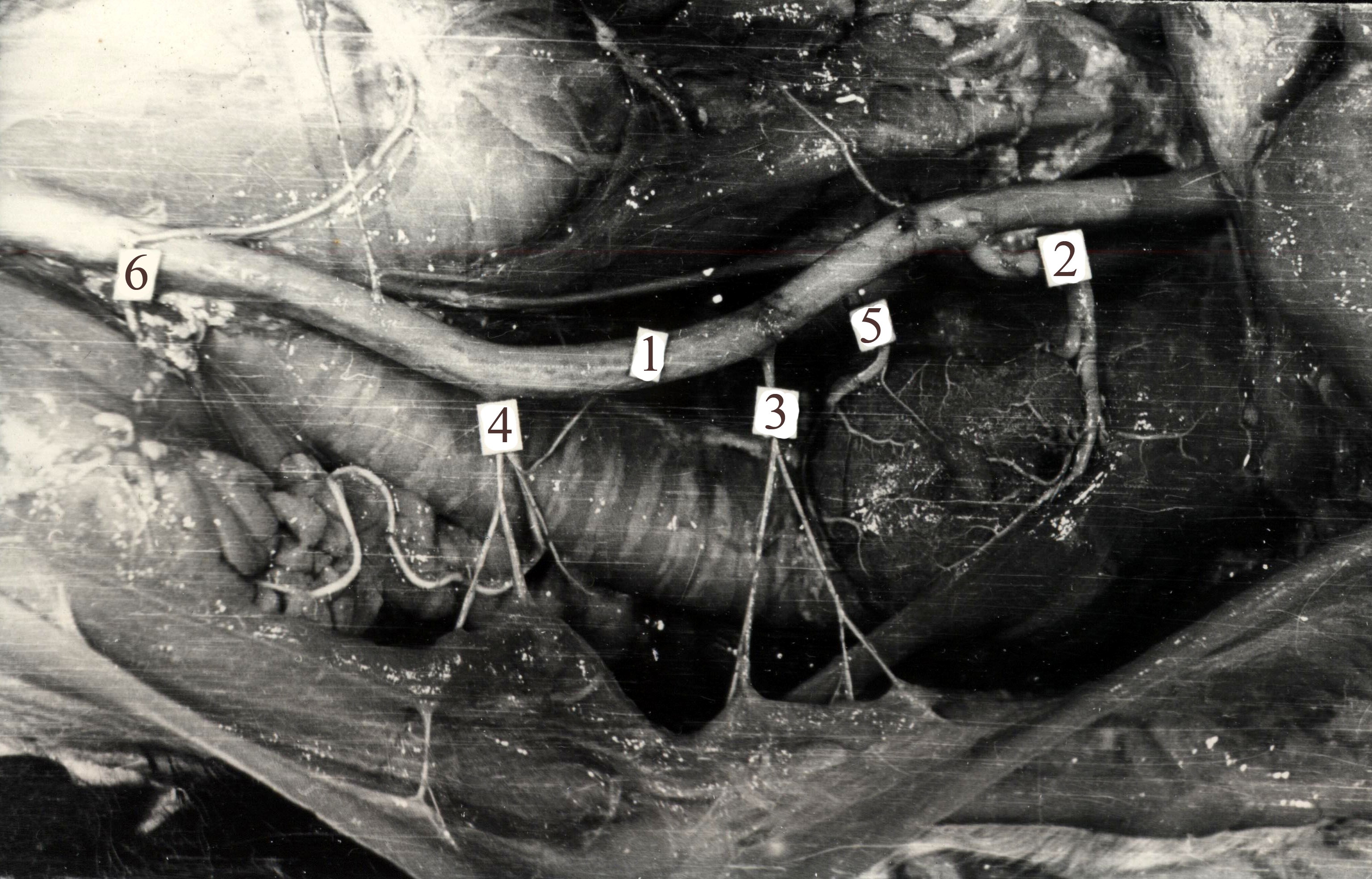
Рис.6. Топография правой общей сонной артерии. 1 – общая сонная; 2 – краниальная щитовидная артерия; 3 – ветви третьего
и 4 – четвёртого шейных позвонков; 5 – каудальная щитовидная артерия; 6 – ветвь пятого шейного позвонка.
Общая сонная артерия ранних плодов имеет прямолинейный ход и просматривается через кожу. Общая картина её ветвления уже соответствует взрослым животным. Отличие заключается в том, что с формированием мышц и органов шеи появляются новые ветви, увеличивается их диаметр, количество
настомозов и частота ячей в сетях, т.е. генезис органов прямо влияет на развитие сосудов, а сосуды – на органы.
Левая и правая общие сонные артерии отдают неодинаковое количество ветвей. Левая 7–9, правая - до 11, диаметром 0,8–1,2 мм. Чаще мышечные ветви отходят общим стволом с ветвями вилочковой железы. Через 1–2,5 см ствол дихотомически делится на дорсальную ветвь, идущую в длинные мышцы шеи, пищевод и вентральную, направляющуюся в трахею, вилочковую железу и в вентральные мышцы шеи. Ветви, диаметром до 2,2 мм, в пищеводно-трахеаль-
ном жёлобе соединяются по типу «конец в конец», образуя коллатераль, которая дополнительно направляет ветви в вентральные мышцы шеи, в пищевод и трахею. В пищеводе, имея рассыпной тип ветвления, образуют сети с крупными и мелкими ячеями полигональной формы.
9. Рост диаметра (мм) общей сонной артерии
Трахеальные ветви, в отличие от пищеводных, распадаются на три – четыре более мелкие составляющие. Ветви образуют сектор кровоснабжения трахеи, включающий 6–8 межкольцевидных пространств. Сеть сектора представлена
крупными ячеями, вытянутыми вдоль трахеальных колец. Сети пищевода, трахеи, мышц и вилочковой железы тесно соединяются между собой.
Диаметр артерии плодов и животных после рождения увеличивается неравномерно. Наиболее активно он возрастает у плодов в шесть и девять месяцев. В постнатальном периоде развития активно калибр артерии увеличивается у телят до шести и молодняка - от 12 до 18 месяцев. Особенно резко диаметр артерии возрастает до 7 дней после рождения. У быков он увеличивается интенсивнее, чем у тёлок. Подобный рост диаметра артерии можно объяснить сменой сред обитания новорожденного и гиперфункцией мышц головы, шеи и всего организма в новорожденный период (табл.9).
Вопросы для контроля
- Какие артерии несут кровь к голове?
- Где располагаются общие сонные артерии?
- Как артерии влияют на развитие мышц, а мышцы на артерии?
- Что образуют сегментальные ветви в пищеводно-трахеальном желобе?
- Как растёт диаметр общих сонных артерий быков и тёлок?
- Как происходит кровоснабжение трахеи и пищевода?
2.6. ОСОБЕННОСТИ РОСТА ГРУДНОЙ И БРЮШНОЙ АОРТЫ
Грудная аорта простирается в средостении от V-VI грудного позвонка до аортального отверстия диафрагмы. По ходу васкуляризирует пищевод, трахею и ножки диафрагмы. С возрастом плодов и животных после рождения диаметр её увеличивается неравномерно и устанавливается возрасте 18 месяцев. В плодном периоде развития диаметр грудной аорты возрастает активнее, чем после рождения, и к рождению достигает 52,97% от диаметра 18-месячных животных.
Абсолютный прирост диаметра грудной аорты активно увеличивается во второй половине плодного периода развития и в 9 месяцев достигает 8 мм, а за 6
месяцев после рождения возрастает только на 5,5 мм и устанавливается в 18 месяцев, затем увеличивается, но незначительно.
В относительных величинах, наоборот интенсивно возрастает от 3 до 6 месяцев (2,77 раза), слабее от 6 до 9 месяцев (1,79 раза), а за 6 месяцев после рождения только в 1,27 раза. До 18-месячного возраста животных диаметр грудной аорты увеличился только на 47,03%. Из этого следует, что диаметр грудной аорты интенсивно возрастает во второй половине плодного периода
развития и до шести месяцев после рождения, и устанавливается в 18 месяцев (табл.10).
10. Рост диаметра грудной аорты
Диаметр брюшной аорты плодов во всех возрастных группах увеличивается достоверно, исключение составляют плоды 7 месяцев. Следует особо отметить активный рост диаметра аорты в девять месяцев, т.е. непосредственно перед рождением.
Абсолютный прирост диаметра брюшной аорты наиболее активно возрастает в четыре, шесть и девять месяцев плодного развития. Если натальный период разделить на первую (3–6) и вторую (6–9 мес.) половины развития, тогда чётко прослеживается следующая закономерность: в первую половину плодного развития абсолютный прирост диаметра брюшной аорты был значительно выше (5,63 мм), чем во вторую (4,66 мм). Чем можно объяснить подобный рост брюшной аорты? Полагаем, что в этот период идёт интенсивное формирование «поля роста» для генезиса зачатков и развития внутренних органов и скелета. К концу беременности и перед рождением сформированные
органы уже функционируют и тот рост диаметра аорты, по которой течёт кровь, достаточен для обеспечения их питательными веществами (табл.11).
11. Рост диаметра (мм.) брюшной аорты
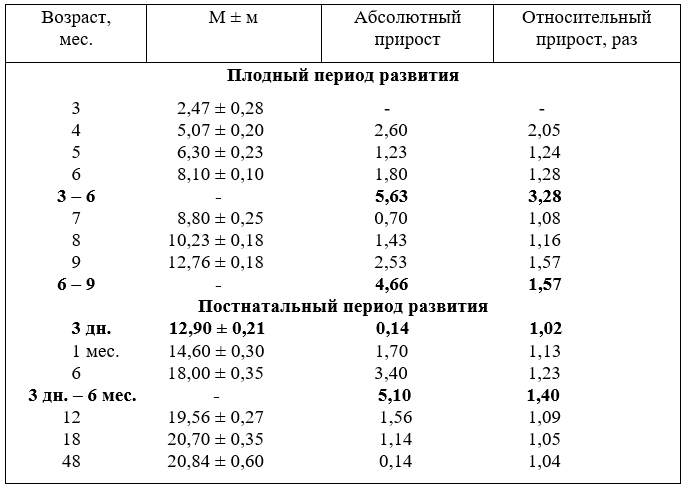
После рождения диаметр брюшной аорты новорожденных и телят в возрасте один, три месяца увеличивается недостоверно, в остальных группах – достоверно. Особенно активно возрастает в шесть месяцев (5,1 мм), что указывает на высокую функциональную активность всех органов, но особенно опорно-двигательного аппарата.
Диаметр брюшной аорты устанавливается в 18 месяцев, затем до четырёх лет увеличивается только на 0,14 мм и отстаёт от абсолютного прироста каудальной полой вены. В целом, среднемесячный прирост диаметра брюшной аорты в плодном периоде развития был выше, чем после рождения.
В относительных величинах наиболее интенсивно диаметр брюшной аорты увеличивается в первую половину плодного развития (3,28 раза) и менее – во вторую (1,57 раза). После рождения, по сравнению с новорожденными, у телят шести месяцев калибр аорты возрастает в 1,4 раза, а в 18 месяцев только в 1,15 раза (табл. 11). В старших возрастных группах диаметр аорты увеличивается на
сотые доли единиц, что существенно не может повлиять на гемодинамику и кровоснабжение органов. Мы их считаем возрастными изменениями.
Брюшная аорта отдаёт комплекс сосудов, снабжающих кровью, как стенки, так и органы брюшной полости. К ним относятся чревная, краниальная брыжеечная, надпочечная, почечная и каудальная брыжеечная артерии и др.
Вопросы для контроля
- Где располагается грудная аорта?
2, Где располагается брюшная аорта?
- Объясните особенности роста грудной?
- Укажите особенности роста брюшной аорты?
- Какие основные артерии отдаёт брюшная аорта?
6, Что кровоснабжает грудная и брюшная аорта?
2.6.1. РОСТ ДИАМЕТРА ЧРЕВНОЙ АРТЕРИИ
Чревная артерия кровоснабжает селезёнку, многокамерный желулок, печень и поджелудочную железу.
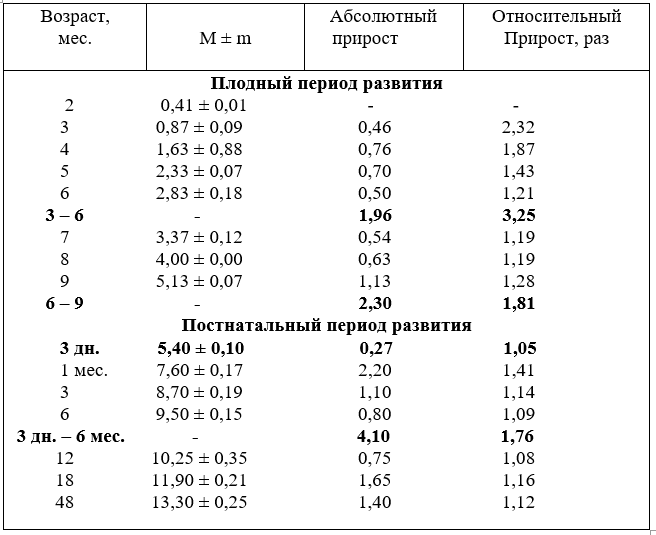
В плодном периоде развития диаметр чревной артерии увеличивается достоверно. Исключение составляют плоды шести и семимесячного возраста. Наиболее активно диаметр артерии увеличивается в три – пять и в девять месяцев.
Наибольший абсолютный прирост диаметра артерии наблюдается также в возрасте плодов четыре, пять и девять месяцев. В первую половину плодного периода развития прирост составил 1,96 мм, а во вторую выше – 2,3 мм.
В относительных величинах активно диаметр артерий увеличивается, наоборот, в первую (3,25) и менее во вторую (1,81 раза).
После рождения калибр чревной артерии увеличивается неравномерно, наиболее активно возрастает у телят в возрасте одного и трёх месяцев и устанав-
ливается в 18 месяцев. До возраста четырёх лет тенденция увеличения диаметра артерии сохраняется и составляет 1,4 мм.
В постнатальном периоде развития наибольший абсолютный прирост диаметра артерии происходит в возрасте телят одного и трёх месяцев. За первое полугодие прирост составил 4,1 мм, что значительно больше, чем во второй половине плодного развития, а в 18 месяцев – 1,65 мм.
В относительных величинах в первую и во вторую половины плодного развития рост диаметра чревной артерии был выше, чем в любом возрасте после рождения. Причём интенсивность роста артерии в плодном периоде к рождению
имеет тенденцию к понижению. Эта тенденция сохраняется и после рождения до четырёхлетнего возраста (табл.12).
12. Рост диаметра (мм) чревной артерии
2.6.2. РОСТ ДИАМЕТРА КРАНИАЛЬНОЙ БРЫЖЕЕЧНОЙ АРТЕРИИ
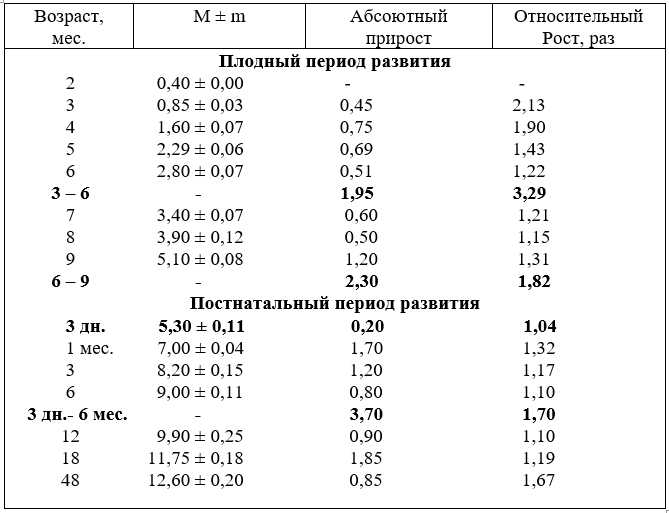
Диаметр артерий достоверно увеличивается во всех возрастных группах плодного периода развития. Наиболее активный рост калибра артерии отмечается у плодов в возрасте четыре, пять и девять месяцев, после рождения у телят в один, шесть и 12, и устанавливается рост диаметра в 18 месяцев, затем к четырём годам увеличивается, но незначительно.
Абсолютный прирост диаметра артерии более активно увеличивается во вторую половину плодного развития (6–9), чем – в первую (3–6 месяцев). После рождения интенсивно калибр артерии возрастает у телят в возрасте один и три месяца, а за первое полугодие после рождения увеличивается на 3,7 мм. То есть, на этот возраст телят приходится пик абсолютного прироста диаметра артерии по сравнению с плодным и первым полугодием постнатального развития (табл.13).
Чёткая закономерность роста сосудов прослеживается из сравнения динамики развития диаметра чревной и краниальной брыжеечной артерий,
13. Рост диаметра (мм) краниальной брыжеечной артерии
обеспечивающие кровоснабжением передний и средний отделы пищеварительной системы. Абсолютный прирост и относительный рост их совпадают, разницу составляют лишь сотые доли единицы (табл.14).
14. Сравнительный рост диаметра (мм) чревной и краниальной брыжеечной артерий
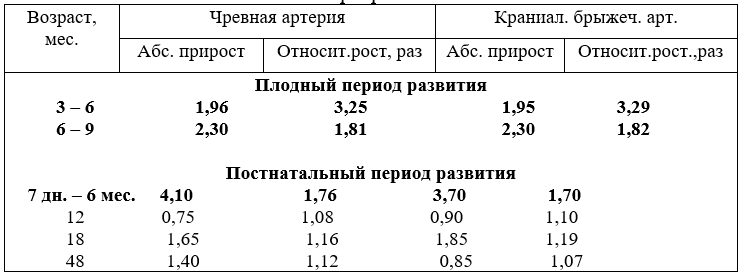
О чём свидетельствует рост этих сосудов? В плодном периоде развития артерии обеспечивают током крови доставку питательных веществ необходимых
для развития, формирования и готовности органов пищеварительной системы к перевариванию пищи постнатальном периоде развития. После рождения молодняк несколько раз меняет род пищи (молозиво, молоко, молоко и грубый корм, затем только грубый корм), Со сменой рода пищи увеличивается площадь пищеварения, всасывания переднего и среднего отделов пищеварительной системы, которые кровоснабжают артерии. Из этого следует, что на каждом этапе развития создаются приспособления органов, систем организма к меняющимся условиям, в которых он живёт, размножается и развивается. Отсюда изменяется диаметр сосудов.
Любой процесс в организме является саморегулирующим. Следовательно: клеткам, тканям, органам и системам на каждом этапе развития соответствует своя форма, строение, среда обитания, обмен веществ и их функция. Для уяснения процессов проходящих в развивающемся организме важно понимание законов биологического саморегулирования.
Некоторые исследователи допускают выражения, что «функция» определяет «материю» и что «функция» всегда предшествует «материи», но без «материи» нет функции, как без «функции» нет «материи». Эти два понятия, на определённом этапе развития, постоянно влияют друг на друга и приводят к саморазвитию органов, в том числе и сердечнососудистой системы.
2.6.3. РОСТ ДИАМЕТРА НАРУЖНОЙ ПОДВЗДОШНОЙ АРТЕРИИ
Наружная подвздошная артерия отходит от брюшной аорты под пятым поясничным позвонком и является основной магистралью для кровоснабжения свободного отдела тазовой конечности. В самом начале она направляет для брюшных стенок окружную глубокую подвздошную артерию, затем в заднебедренную группу мышц – глубокую бедренную и переходит в бедренную артерию, опускающуюся дистально до копытец конечности.
Диаметр наружной подвздошной артерии с возрастом плодов увеличивается достоверно и с возрастающей активностью. Наиболее интенсивно он возрастает в
шесть и девять месяцев плодного периода развития и у взрослых животных достигает 14,6 мм табл.15).
Абсолютный прирост диаметра артерии увеличивается наиболее активно в возрасте плодов шесть, семь и девять месяцев. В первую половину плодного
15. Рост диаметра (мм) наружной подвздошной артерии
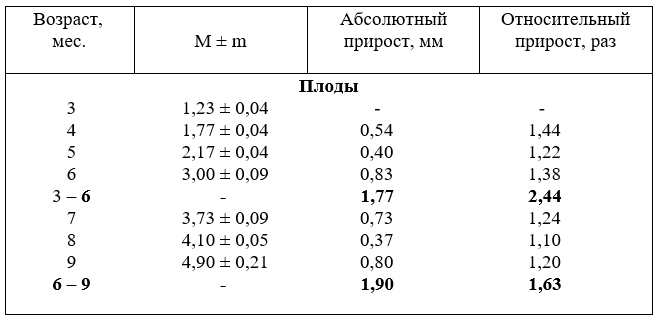
развития абсолютный прирост ниже (1,77 мм), а во вторую выше (1,90 мм). В относительных величинах, наоборот, рост диаметра происходит активнее в первую половину плодного развития (2,44 раза), чем во вторую (1,63 раза).
Причём, разница существенная (0,81 раза).
2.6.4 РОСТ ДИАМЕТРА ВНУТРЕННЕЙ ПОДВЗДОШНОЙ АРТЕРИИ
Брюшная аорта, отдав в задние конечности правую и левую наружные подвздошные артерии, переходит назад в общий ствол внутренних подвздошных артерий. У крупного рогатого скота общий ствол артерий в области шестого поясничного позвонка разветвляется на правую и левую внутренние подвздошные артерии, у плодов диаметром 0,67 – 3,53 мм, у взрослых животных достигает 7,6 мм. Они являются основной магистралью для кровоснабжения стенок и органов тазовой полости. Артерии в области малой седалищной вырезки отдают задние ягодичные артерии и переходят во внутренние срамные.
На своём пути она направляет париетальные ветви в стенки таза и висцеральные – в органы тазовой полости.
Диаметр внутренних подвздошных артерий в плодном периоде развития увеличивается неравномерно и по сравнению с другими сосудами замедленно, как бы он отстаёт в развитии, но к рождению темп роста его всё – таки увеличивается. Особенно интенсивно он возрастает после рождения до года, а
затем темп роста его замедляется и устанавливается в 18 месяцев (табл.16).
16. Рост диаметра (мм) внутренних подвздошных артерий
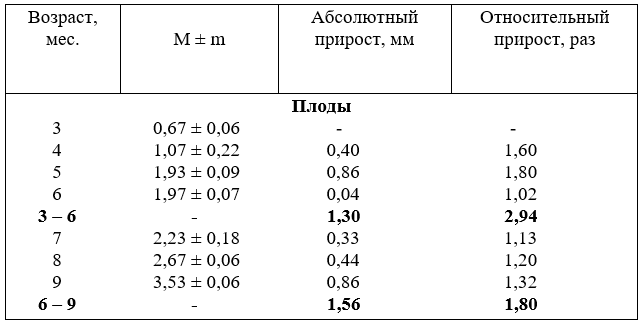
Абсолютный прирост диаметра наиболее активно возрастает у плодов в пять и девять месяцев и за первую половину плодного развития он увеличился только на 1,3 мм, а за вторую – на 1,56 мм, а за весь плодный период – на 2,86 мм. После
рождения до полутора лет на 4,07 мм.
В относительных величинах наиболее активно возрастает диаметр артерии в первой половине плодного развития – в 2,94 раза, а за вторую – только в 1,8 раза,
а за весь плодный период – 5,26 раза. Для сравнения, до 18 месячного возраста увеличивается только в 2,15 раза.
Вопросы для контроля
- Перечислите особенности роста диаметра чревной артерии?
- Когда устанавливается роста диаметра чревной артерии?
- Какие особенности роста краниальной брыжеечной артерии?
- Как влияет или нет смена рода пищи на рост сосудов и площадь всасывающей поверхности кишечника?
- Может функция быть без материи или материя без функции?
- На какие конечные сосуды делится брюшная аорта?
- Какие органы снабжает кровью наружная подвздошная артерия?
- Какие органы снабжает кровью внутренняя подвздошная артерия?
- В какую артерию переходит внутренняя подвздошная артерия?
3. МОРФОФУНКЦИОНАЛЬНОЕ ОБОСНОВАНИЕ ВОЗРАСТНЫХ ИЗМЕНЕНИЙ ДУГИ АОРТЫ И ЕЁ АРТЕРИЙ
Описанные сегменты дуги аорты с возрастом животных увеличивают диаметр неравномерно как в целом, составляющих дугу, так и каждый сегмент
отдельно. Особенно большим колебаниям в росте подвержен второй сегмент относительно третьего и артериального протока. Из приведённого анализа следует, что артериальный проток относительно третьего сегмента дуги аорты плодов выполняет большую функциональную нагрузку, чем второй сегмент.
Наименьшие размеры второго сегмента в плодном периоде развития, способствуют местному сопротивлению движения крови из левого желудочка сердца в грудную аорту. Сформировавшаяся, таким образом, «горловина», располагающаяся на месте второго сегмента дуги аорты, увеличивает кровоток в плечеголовной ствол плода. Что способствует развитию головного мозга и опережающему росту передней части тела плода, относительно задней.
Одновременно поток крови, идущий через артериальный проток в третий сегмент дуги аорты, способствует оттоку крови из первого сегмента в третий через «горловину» по принципу инжектора (см. Чугаев Р.Р. Гидравлика: закон Бернулли – 1970, с.85–91). Отсюда следует, что второй сегмент в плодном периоде развития, выполняет функцию анастомоза между первым и третьим сегментами дуги аорты. После облитерации артериального протока в возрасте телят 1 – 1,5 месяца, диаметр его выравнивается относительно первого и третьего
сегментов и начинает выполнять основную функцию проведения крови из левого желудочка в грудную аорту. В результате задняя часть тела к этому возрасту выравнивается в развитии по массе и росту с передней.
Диаметр плечеголовного ствола возрастает более равномерно, чем сегменты дуги аорты, и стабилизируется в 18 месяцев постнатального периода развития. Установлено, что внешний диаметр ствола и его внутренний калибр увеличиваются относительно друг друга неодинаково и связано это с неравномерностью роста его толщины стенок. Неравномерность роста толщины стенок объясняется возрастающим сопротивлением току крови к развивающимся органам передней части тела, что приводит к развитию мышечной оболочки стенок сосудов. У быков она активнее развивается до 18 месяцев, чем у тёлок.
Диаметр подключичных артерий был самым широким (62,2%), по отношению к плечеголовному стволу, в возрасте плодов трёх месяцев. Затем рост диаметра к рождению и после рождения до четырёх месяцев несколько
замедлялся. Поэтому самыми узкими относительно к стволу (39,9%) артерии были у телят в возрасте шести месяцев после рождения. К 18-месячному возрасту диаметр их вновь увеличивался (54,7%). Такое увеличение диаметра подключичных артерий связано с повышением функциональной нагрузки на грудные стенки, конечности и мышцы шеи животного.
Диаметр ствола сонных артерий значительно превосходит диаметр подключичных артерий. Так, в плодном периоде развития калибр его был выше диаметра подключичных артерий в 1,49 раза, а в постнатальном - в 1,34 раза. Остаётся самым широким в возрасте плодов три месяца (95,7%). К рождению он понижается до 73,7% и на этом уровне сохраняется до трёх месяцев после рождения. Затем наступает период его волнообразного роста и спада, который стабилизируется в 18 месяцев (79,8%). Поэтому, во всех возрастных группах плодного и постнатального периодов развития основным сосудом, являющимся продолжением плечеголовного ствола, следует считать ствол сонных артерий.
Диаметр общих сонных артерий увеличивается неравномерно до 18 месяцев. Особенно калибр артерий активно увеличивается у телят от трёх дней до одного месяца после рождения. У быков он возрастает более интенсивно, чем у тёлок и разница в диспропорции их роста устанавливается в 12 месяцев.
Все артерии шеи имеют магистральный тип ветвления и образуют внутрисистемные и межсистемные анастомозы и коллатерали. Внутрисистемные
анастомозы в фасциях и наружном перимизии мышц образуют более грубые, сети, внутри мышц – тонкие, ячеи которых вытянуты по ходу мышечных волокон и пучков.
Вопросы для контроля
- Чем объяснить опережающий рост передней части тела плода?
- В каком возрасте происходит облитерация артериального протока?
- Чем объяснить неравномерность роста диаметра плечеголовного ствола?
- Чем объяснить неравномерность роста диаметра подключичных артерий?
- Какой сосуд следует считать продолжением плечеголовного ствола и почему?
- В каком возрасте между быками и тёлками устанавливается разница в росте диаметра общего ствола сонных артерий?
- Какие анастомозы и коллатерали устанавливаются в области шеи и головы?
4. МОРФОФУНКЦИОНАЛЬНОЕ ОБОСНОВАНИЕ РОСТА ГРУДНОЙ И БРЮШНОЙ АОРТЫ
Из сравнения абсолютного прироста диаметра грудной относительно к брюшной аорте (табл.17) следует отметить, что диаметр грудной аорты на уровне V–VI грудного позвонка активнее возрастает, чем брюшной аорты на уровне ХIII грудного и первого поясничного позвонка. Это соотношение в первую половину плодного периода развития составляет как 1:1,4, а во вторую - как 1:1,7 раза, что значительно выше, чем до шести месяцев. К возрасту телят шесть месяцев после рождения эта диспропорция в росте понижается и составляет как 1:1,08.
17. Абсолютный прирост и относительный рост диаметра грудной и брюшной аорты
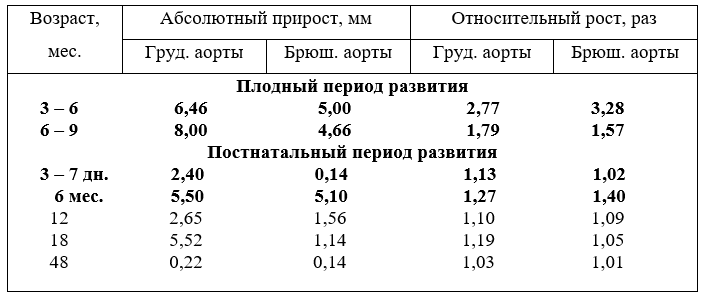
В абсолютных величинах прирост диаметра грудной и брюшной аорты активно увеличивается от шести до девяти месяцев плодного развития и несколько ниже от 3 до 6. После рождения до возраста шести месяцев диаметр грудной и брюшной аорты возрастает почти одинаково. И всё же диаметр грудной аорты продолжает активнее увеличиваться до 18 месяцев, чем брюшной. Считаем, что на прирост диаметра грудной аорты влияет повышенная динамическая нагрузка, приходящая на мышцы передней части тела, особенно, на мышцы плечевого пояса, шеи и на вдыхатели, и выдыхатели.
В относительных величинах рост диаметра грудной и брюшной аорты активно возрастает до 6 месяцев плодного периода развития, к рождению и после
рождения до 6 месяцев телят плавно понижается и устанавливается в 18 месяцев. До четырёх лет калибр их возрастает, но незначительно (табл.17). У крупного
рогатого скота увеличение диаметра сосудов идёт более равномерно, с некоторым подъёмом в пять, семь и девять месяцев плодного периода развития, а также в 3 – 7 дней и в шесть месяцев после родов.
Вопросы для контроля
- Чем объяснить активный рост диаметра грудной аорты относительно брюшной?
- В каком возрасте устанавливается рост грудной и брюшной аорты?
5. АБСОЛЮТНЫЙ ПРИРОСТ И ОТНОСИТЕЛЬНЫЙ РОСТ АРТЕРИЙ ПЛЕЧЕГОЛОВНОГО СТВОЛА
Диаметр третьего сегмента дуги аорты по сравнению с другими артериями наиболее активно приростает в первую и вторую половины плодного периода развития, затем плечеголовной ствол, общий ствол сонных артерий, общие сонные артерии и последние - подключичные (табл. 18).
18.Абсолютный прирост диаметра (мм) артерий дуги аорты
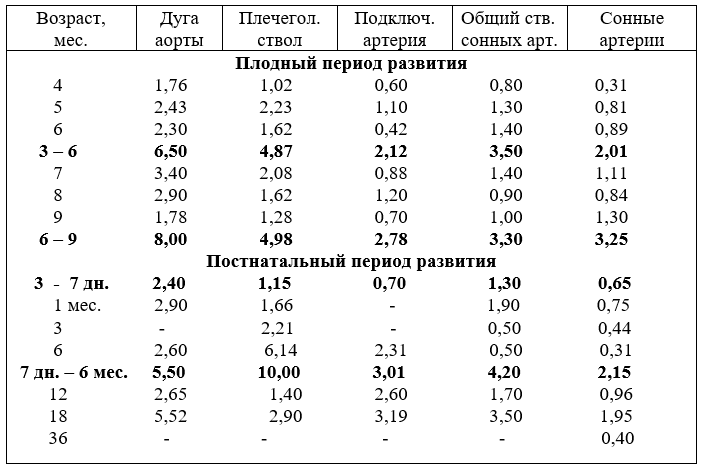
Из таблицы видно, что прирост диаметра плечеголовного ствола - основного сосуда, снабжающего кровью переднюю часть тела, в плодном периоде развития отстаёт в приросте от калибра дуги аорты, которая током крови питает заднюю часть тела. Такой порядок прироста диаметра артерий сохраняется до возраста
одного месяца телят после рождения, что выравнивает рост задней части тела относительно передней.
В шесть месяцев после рождения порядок абсолютного прироста калибра сосудов меняется. На первое место выступает плечеголовной ствол, на второе – грудная аорта, затем идёт общий ствол сонных артерий и, наконец, подключичные артерии и на последнем месте располагаются общие сонные артерии. В старших возрастных группах прирост сосудов соответствует порядку, приведенному в плодном периоде развития. Становится вопрос – чем объяснить перманентный абсолютный прирост калибра артерий? Считаем, сменой среды обитания, мышечной гиперфункцией и продолжением формирования передней части тела, особенно, головы и головного мозга, грудных конечностей.
19. Относительный рост диаметра артерий дуги аорты
Относительные, величины роста диаметра сосудов грудной аорты и плечеголовного ствола показывают, что активнее калибр всех артерий возрастает в первую половину плодного развития и слабее – во вторую. Но темп роста плечеголовного ствола, значительно выше, чем остальных сосудов. Во второй половине плодного периода развития, к рождению, по относительному росту
диаметра сосуды расположились в следующем порядке: общие сонные артерии, плечеголовной ствол, подключичные и общий ствол сонных артерий (табл.19).
Но у телят в шесть месяцев после рождения на первое место выходит плечеголовной ствол, на второе – подключичные артерии, затем дуга аорты, общие сонные артерии и последним ствол сонных артерий. Что свидетельствует о воздействии на развитие сосудов как смены среды обитания, так и гипердина-
мии и перехода питания телят от молозива, молока к растительной пище. Абсолютный прирост и относительный рост сосудов устанавливается в возрасте 18 месяцев.
Вопросы для контроля
1. Как увеличивается диаметр плечеголовного ствола относительно дуги аорты у телят до одного месяца?
2. Как меняется порядок абсолютного прироста диаметра дуги аорты и плечего- головного ствола в возрасте телят шесть месяцев?
6. АБСОЛЮТНЫЙ ПРИРОСТ И ОТНОСИТЕЛЬНЫЙ РОСТ АРТЕРИЙ БРЮШНОЙ АОРТЫ
В первой половине плодного периода развития абсолютный прирост диаметра артерий брюшной аорты происходит неравномерно, активно он увели-
чивается в четырёхмесячной возрастной группе плодов, в остальных значительно ниже, а во второй – в девять месяцев. Причём, в возрасте плодов девять месяцев среднемесячный прирост калибра артерий выше, чем в любой другой возрастной группе плодов, но прирост диаметра брюшной аорты за первую половину плодного развития выше на 0,97 мм, чем за вторую, а у остальных ниже (табл. 20).
20. Абсолютный прирост диаметра (мм) артерий брюшной аорты
Компенсация в росте абсолютного прироста диаметра брюшной аорты происходит после рождения, особенно, у молодняка с одного до шестимесячного возраста и достигает 5,1 мм, чревной и краниальной брыжеечной артерий, соответственно, 4,37 и 3,90 мм. Высокий темп прироста калибра артерий сохраняется до возраста 18 месяцев и устанавливается в два года. Эти величины абсолютного прироста диаметра брюшной аорты убедительно показывают, что у телят до трёх месяцев, т.е. в молозивный и молочный периоды питания, калибр
артерий увеличивается замедленнее, чем в период питания растительной пищей, которая активирует абсолютный прирост калибра всех сосудов.
Н.А.Малков (1983) пишет, что в постнатальном периоде развития минимальный среднемесячный прирост диаметра большинства артерий наблюдается у телят после рождения, что связано со сменой среды обитания. В этом случае повышенная функциональная активность органов и систем требует повышенного притока крови к ним с кислородом и питательными веществами. На что сосуды отвечают ростом диаметра, а отсюда притока большего объёма крови с питательными веществами.
После рождения активный относительный рост продолжается до возраста телят шести месяцев. Если в относительных величинах рассматривать совместно плодный и постнатальный периоды развития, тогда проявляется следующая
закономерность, чем старше по возрасту плод, животное после рождения, тем ниже относительный рост (табл. 21).
21. Относительный рост брюшной аорты и её дериваций
Общеизвестно, что относительные величины более точно указывают на развитие кровеносной системы. Брюшная аорта и её деривации в первую половину плодного периода развития увеличивают диаметр активнее, чем во вторую половину.
В научной литературе имеются сведения о морфофункциональном состоянии артерий (Б.П.Шевченко, 1984, 1999; Х.Б. Баймишев, И.В. Хрусталёва, 2001; А.Гончаров, 2002;.М.С.Сеитов, 2002), указывающие на перманентность роста и абсолютного прироста диаметра артерий как в плодном, так и в постнатальном периодах развития. Смена периодов показывает, что абсолютный прирост диаметра сосудов выше во вторую половину плодного развития и ниже – в первую.
После рождения, в связи с особенностями внешней среды (экологии), смены рода пищи, повышенной динамической нагрузки продолжается активный рост диаметра артерий до шести месяцев, затем стабилизируется в 18 – 24 месяца. Но фактический материал, характеризующий динамику роста сосудов в возрастном аспекте, ещё беден. Он не позволяет сделать количественные и качественные (морфометрические) обобщения.
Изучение сердечнососудистой системы с возрастом имеет не только теоретическое, но и важное практическое значение как указывали ведущие учёные (Ф.Н. Валькер, 1954; Б.П.Токин, 1970; Г.М.Удовин, 1972; О.В.Алексеев, 1981) и др.
Вопросы для контроля
1. Как с возрастом меняется абсолютный прирост диаметра брюшной аорты?
2. Как и почему меняется прирост диаметра брюшной аорты и артерий после рождения?
3. Как с возрастом животных изменяется относительный рост диаметра сосудов в относительных величинах?
7. ВЕНЫ
Вены (vena – кровеносный сосуд, жила) выходят из венозного звена микроциркуляторного русла и несут кровь насыщенную двуокисью углерода, продуктами метаболизма, гормонами, питательными веществами, кислородом в правое предсердие и к органам выделения. Объединяясь, они образуют ёмкостные сосуды – краниальную и каудальную полые вены, вены сердца, открывающиеся в венозный синус правого предсердия, лёгких.
Передняя полая вена выносит кровь из передних конечностей, грудных стенок, шеи и головы, задняя – из тазовых конечностей, органов тазовой полости, брюшных стенок. Из органов брюшной полости кровь оттекает в воротную вену печени, а затем в каудальную полую вену. Из этого следует, что печень стоит на пути оттока крови от органов пищеварительной системы и селезёнки.
Кровь из позвоночного столба человека собирается в венозный коллектор, из которого далее оттекает в верхнюю и нижнюю полые вены (В.Я.Протасов, 1961). У крупного рогатого скота и северного оленя лобная, поверхностная, общая и глубокая лицевые вены, язычный лицевой венозный ствол, надглазничная, венозное сплетение глазного яблока, поверхностная височная и верхняя челюстная вена формируют коллектор головы, из которого назад выходит наружная ярёмная вена (Б.П.Шевченко, 1975).
Правый и левый коллекторы позвоночного столба и головы соединяются между собой посредством поперечных венозных анастомозов, сетей, сплетений и
кавернозных тел носа, носоглотки, шеи, спинного мозга и половых органов тазовой полости, формируя венозный комплекс сосудов (М.В.Андреев, 1988).
Часто вены в местах изгибов, мышечных динамических нагрузок, пережатий образуют расширения в виде ампул, а между ними сужения. В листках фасций, серозных оболочек, капсул, иногда в стенках трубчатых органов, мочевого пузыря одну артерию могут сопровождать две вены, соединяющиеся между собой посредством поперечных вено-венулярных, артериоло-венулярных анастомозов. При слиянии вены образуют соустья, стоки, каверны, синусы. Соустья вен отделяют мысы, которые предотвращают гемодинамические столкновения, сливающихся потоков крови (рис.7).

Рис.7. Слепок венозных сплетений преддверия носа. Латекс. Об. 3,5; ок. 5.
Стоки, синусы чаще образуют вены головного мозга, трубчатых и губчатых костей, но их много в кавернозных телах слизистой оболочки носа, носоглотки, половых органов. Кроме этого в кавернозных телах встречаются «озёра», а в ампулах вен встречаются трабекулы (рис.8).

Рис. 8. Каверна. Импрегнация азотнокислым серебром. Об.40, ок. 15.
В просвете вен встречаются полулунные пластинки, складки, одно, двух, трёх и более створчатые клапаны, гребни, трабекулы, лакуны. Пластинки мы рассматриваем как остатки венозных клапанов (рис.9).
Рис. 9. Продольный разрез вены, видны клапаны, складки
и другие внутривенные образования.
Складки чаще встречаются в просвете изгибов вен, гребни – в точке прилежания вены к костной основе, трабекулы – в ампулообразных расширениях лицевых, костных, иногда в наружных ярёмных венах, лакуны – в точке бывшего клапана вены или фиксации к костной основе. Комплекс внутривенных образований выполняет тонкую гемодинамическую функцию, особенно, мысы, складки, гребни, которые чаще располагаются у соустье притоковых вен.
Трабекулы укрепляют стенку вены от чрезмерного растяжения и надрыва, поэтому они чаще располагаются в местах компрессионных нагрузок. Их делят на тонкие, средние и толстые.
Ампулообразные расширения вен, сетей, сплетений и кавернозных тел разделяют сужения или синусы, каверны и т.д. В стенке расширений гладкие мышечные клетки располагаются разреженно, а в сужениях – чаще или плотнее.
В стенке отводящих вен слизистой оболочки носа, кавернозных тел и сплетений глаз встречаются жомы, состоящие из двух слоёв мышечных клеток, расположенных относительно друг друга перпендикулярно. К ним подходят нервные волокна. Жомы, перекрывая отток крови из ампулообразных расширений, способствуют переполнению их кровью и усиливают механизм эрректильного действия венозных сетей, сплетений, кавернозных тел. Особенно
в слизистой оболочке носа, вследствие чего, перекрываются его дыхательные ходы. Задача врача при рините открыть эти жомы (рис.10).

Рис. 10. Жом вены. Импрегнация азотнокислым серебром.
Об. 20, ок. 10.
Ёмкость венозного русла в норме составляет 39–55%, в отдельных случаях может достигать 65% и превосходит содержание крови в артериях в 1,8–2,2 раза
Роль вен заключается в том, что они выносят кровь насыщенную двуокисью углерода, продуктами обратного обмена, гормонами через воротную систему в общий круг кровообращения. Все вены, кроме легочных и пупочных, несут венозную кровь.
В конечностях различают поверхностную и глубокую венозные магистрали. Поверхностные магистрали формируются плоскими венозными сетями кожи, листков поверхностной фасции и их стволы не сопровождают артерии, глубокие магистрали идут вместе с артериями, нервами, формируя триады и, в соответствии с их делениями, отдают дериваты.
Вопросы для контроля
- Из какого звена микроциркуляторного русла выходят вены?
- Куда по венам течёт кровь?
- Куда открываются передняя и задняя полые вены?
- Из каких частей тела выносят кровь передняя и задняя полые вены?
- Что формируют вены головы?
- Откуда выходит наружная ярёмная вена?
- Какие внутривенные образования встречаются в просвете вен и их функция?
- Какую функцию выполняют «жомы» в спелетения и кавернозых телах?
- Перечислите внутривенные образования и укажите их функцию?
- Какие венозные магистрали различают в конечностях?
8. СИСТЕМА КРАНИАЛЬНОЙ ПОЛОЙ ВЕНЫ
Передняя полая вена отводит кровь в правое предсердие сердца из головы, шеи, грудных стенок и конечностей, образована слиянием плечеголовной вены с подключичными венами. В систему входят наружные и внутренние ярёмные, позвоночные, глубокие шейные, дорсальные лопаточные, подключичные и внутренние грудные вены. Описание вен в монографии дано с периферии тела, т.е. по току крови и слиянию вен к сердцу (по принципу конвергенции), что считаем правильным, а не от сердца против тока крови как дано в руководствах.
8.1. РОСТ ДИАМЕТРА НАРУЖНОЙ ЯРЁМНОЙ ВЕНЫ
У крупного рогатого скота наружная ярёмная вена, диаметром 2,0–24, 0 мм, располагается в ярёмном жёлобе между листками поверхностной фасции, прикрытая платизмой и подкожной мышцей шеи. В просвете вены плодов шести – семи месяцев находится до трёх двустворчатых клапанов, лежащих через 5,0–5,5-3,5 см. Непосредственно перед рождением количество их в просвете вены увеличивается до пяти, из которых первые три, как правило, трёхстворчатые, а последующие два – двухстворчатые. Расстояние между ними, соответственно равно 2,0; 6,5; 7,0 и 2,0 см. После рождения количество клапанов увеличивается до семи. Все они переходят в трёхстворчатые. Обычно первый клапан в просвете вены располагается на уровне заднего края краниальной щитовидной железы, второй – после него. В средней части наружной ярёмной вены клапаны лежат на уровне шейных позвонков, на расстоянии 5,0–9,5 см. Поэтому наиболее безопасным местом для взятия крови на исследование без повреждения клапанов следует считать участок вены между третьим и пятым шейными позвонками.
Из анализа расположения клапанов в просвете наружной ярёмной вены следует, что с ростом в длину шеи и вены возрастает количество клапанов, а с увеличением диаметра – количество створок в них.
Диаметр наружной ярёмной вены достоверно увеличивается до трёх лет. Наиболее интенсивно он возрастает у плодов шести – семи месяцев, а после рождения у молодняка в один и три месяца (табл.22). В вену открываются затылочная, краниальная щитовидная вена, ветви околоушной, нижнечелюстной железы, мышечные, трахеальные и пищеводные ветви (А.Г.Гончаров,2000).
22. Рост диаметра наружной ярёмной вены
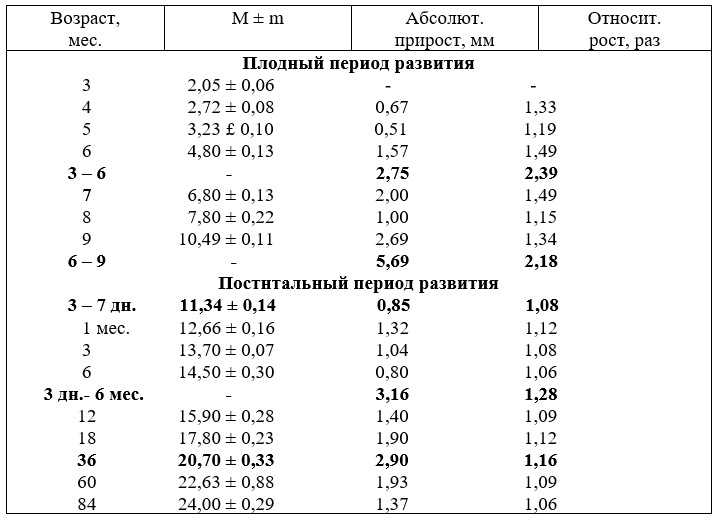
Абсолютный прирост диаметра наружной ярёмной вены активно увелчивает-
ся у плодов в возрасте шесть, семь и девять месяцев. В первую половину плодного периода развития он был значительно ниже (на 3,65 мм), чем во вторую. После рождения диаметр вены возрастает до одного месяца, а у молод-
няка до шести месяцев всего на 2,75 мм. Особенно активно идёт прирост диаметра вены от шести месяцев до трёх лет (на 6,2 мм). К семи годам прирост диаметра вены плавно понижается.
В относительных величинах активно диаметр вены возрастает в первую половину плодного периода развития (2,39 раза), ниже – во вторую (2,18 раза), а за шесть месяцев постнатальной жизни у молодняка увеличивается только в 1,28, а за три года – в 1,16 раза, т.е с шести месяцев до трёх лет прирост остаётся на одном уровне, затем плавно снижается
Из этого следует, что в абсолютных величинах прирост диаметра вены выше во вторую половину плодного периода развития и значительно ниже в первую. После рождения калибр вены максимально возрастает у молодняка до шести месяцев и сохраняет высокий темп роста до трёх лет. Затем в старших группах
плавно понижается. В относительных величинах, наоборот, наиболее активно возрастает в первую и ниже – во вторую половины плодного развития, после рождения низким сохраняется у молодняка до шести месяцев и устанавливается в три года.
А.М.Геселевич (1960) отмечает, что вопросы практики и теории требуют изучения диаметра приносящих и отводящих сосудов. В наши дни анализ соотношения диаметров рядом расположенных артерий и вен как способ оценки функционального состояния венозного русла ещё не решён. Сравнение притока крови к функционирующему органу и оттока от него всегда отражает изменение функции и его кровоснабжения, пишет А.И. Никоноров (1970).
На величину диаметра и толщину стенок сосудов влияют возраст, пол и среда (П.Г.Светлов, 1978). Диаметр артерий женских индивидумов меньше, чем у мужских. С возрастом увеличивается диаметр артерий и вен. Все наблюдаемые изменчивости представляют, в конечном счёте, реакцию развивающегося организма или его частей на изменение условий среды. Сюда следует добавить и функциональной нагрузки.
8.2. Особенности топографии внутренней ярёмной вены
Внутренняя ярёмная вена имеется не у всех домашних животных. У крупного рогатого скота она встречалась в 37-43% случаев, в остальных - отсутствует. У взрослых животных, при наличии, диаметр её достигает 6,2 мм. Располагается она в жёлобе между пищеводом и трахей, медиальнее от общей сонной артерии и
вагуса. Вена выходит из слияния притоковых ветвей венозных сетей глотки, гортани, пищевода, трахеи, мышц шеи, вилочковой железы. В каудальной части шеи она открывается в наружную ярёмную вену, образуя общий ствол ярёмных вен. Правый и левый стволы, сливаясь, образуют плечеголовную вену, в которую открываются подкожные головные, подключичные вены, затем она переходит в краниальную полую вену.
8.3. РЁБЕРНО-ШЕЙНАЯ ВЕНА
Реберно-шейная вена домашних животных выходит из слияния позвоночной,
глубокой шейной, дорсальной лопаточной и первой межреберной вен. В соустье с краниальной полой веной располагается трёхстворчатый клапан.
8.3.1. РОСТ ДИАМЕТРА ПОЗВОНОЧНОЙ ВЕНЫ
Позвоночная вена крупного рогатого скота, диаметром 1,14–6,33 мм, образуется слиянием межпозвоночных вен и притоковых ветвей из дорсальных мышц шеи. Неравномерный, но интенсивно диаметр вены увеличивается у плодов перед рождением и в первый месяц после рождения, особенно до семи дней и достоверно возрастает до 18-месячного возраста. В соустье с глубокой шейной веной находится трёхстворчатый клапан (табл.23).
23. Рост диаметра позвоночной вены
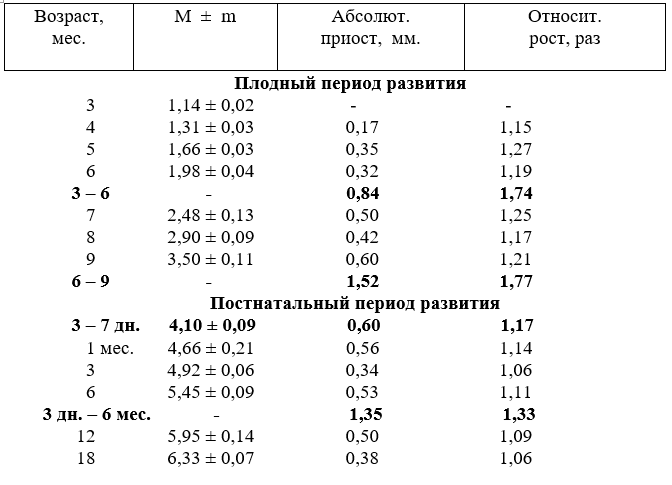
Абсолютный прирост диаметра позвоночной вены в плодном периоде развития происходит волнообразно и всё же к рождению достигает максимальных величин (0,6 мм), но в первую половину внутриутробного развития был ниже, чем во вторую Особенно активно прирост диаметра вены происходит у новорожденных и у телят молочников до одного месяца (1,16 мм).
В относительных величинах, в любой возрастной группе, интенсивнее диаметр развивается в плодном развитии и слабее – после рождения. Если за время внутриутробного развития диаметр вены увеличился в 3,1 раза, то после рождения до возраста 18 месяцев только в 1,6 раза. Из этого сравнения хорошо
видно, что на рост калибра вены резко влияет смена среды обитания и повышенная функция мышц шеи после рождения.
8.3.2. РОСТ ДИАМЕТРА ГЛУБОКОЙ ШЕЙНОЙ ВЕНЫ
Вена выносит кровь из глубоких слоёв дорсальных мышц шеи и головы, между шестым и седьмым шейными позвонками открывается в позвоночную вену. В её соустье находится, как правило, двух или трёхстворчатый клапан. Диаметр глубокой шейной вены у взрослых животных достигает 6,22 мм, достоверно увеличивается до 12 месяцев после рождения и стабилизируется в росте в 18 месяцев, затем возрастает, но незначительно (табл. 24).
24. Рост диаметра глубокой шейной вены
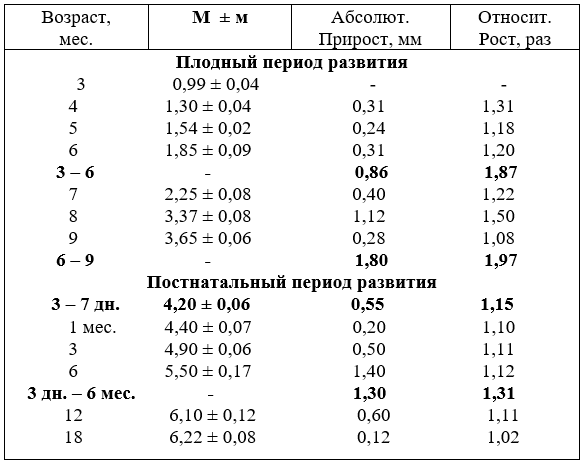
Абсолютный прирост диаметра вены в первой половине плодного периода развития был значительно ниже, чем во второй. Если рассматривать абсолютный прирост калибра вены по возрастным группам плодов, просматривается следующая закономерность, что он неравномерно увеличивается до восьмимесячного возраста и к рождению понижается.
Несколько иная закономерность прироста диаметра вены в постнатальном развитии. Максимальный абсолютный прирост калибра вены наблюдается в новорожденном возрасте и у телят в шесть месяцев после рождения. В первой
половине года он был значительно выше, чем во втором – от шести до 12 месяцев и, затем постепенно понижается до 36 месяцев.
В относительных величинах рост диаметра глубокой шейной вены в первой половине плодного развития был несколько ниже, чем во второй, и начиная с новорожденного возраста до шести месяцев после рождения постепенно понижается, максимума достигает в возрасте телят шесть месяцев. Из данного анализа следует, что рост диаметра вены с возрастом животных ослабевает и устанавливается в 18 месяцев.
8.3.3. РОСТ ДИАМЕТРА ПОДКЛЮЧИЧНОЙ ВЕНЫ
Подключичная вена крупного рогатого скота, диаметром 1,75–17,61 мм, открывается на границе перехода плечеголовной вены в краниальную полую вену. В соустье её находится двух или трёхстворчатый клапан.
Диаметр вены достоверно увеличивается до 18 месяцев постнатального периода развития. Наиболее активно он возрастает у плодов в восемь и девять месяцев, и после рождения - у новорожденных, и телят до шести месяцев. Хотя рост диаметра вены в 18 месяцев достоверный, но он значительно уступает скорости роста его до года.
В абсолютных величинах прирост диаметра подключичной вены, в целом, активно возрастает в плодном периоде развития, причём во второй половине интенсивнее, чем – в первой на 3,11 мм. После рождения активный прирост диаметра вены продолжается до года, но у телят в шесть месяцев был выше на - 2,77 мм, чем – до 12 месяцев и устанавливается в 18 месяцев.
Относительные величины, наоборот, показывают, что интенсивный рост подключичной вены активно возрастает в первой половине плодного периода развития и слабее – во второй и ещё слабее у телят до шести месяцев после рождения. Причём активность роста поперечника вены показывает на постоянно понижающий характер роста в относительных величинах в возрастном аспекте (2,53; 2,31; 1,32 раза, табл. 25).
25. Рост диаметра подключичной вены
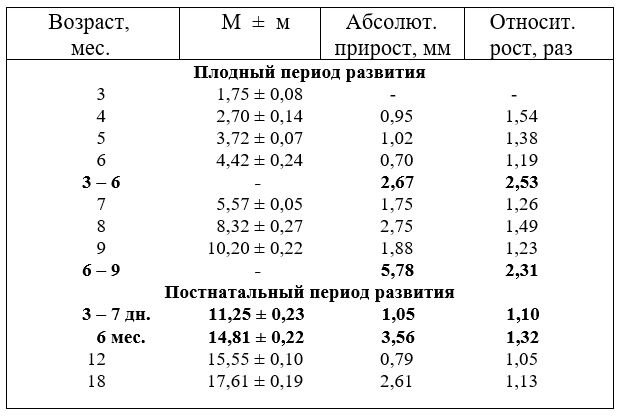
8.3.4.ОСОБЕННОСТИ РОСТА КРАНИАЛЬНОЙ ПОЛОЙ ВЕНЫ
Краниальная полая вена крупного рогатого скота, диаметром 2,8 – 31,6 мм, у плодов и молодняка проходит в грудную полость под дыхательным горлом, через грудную долю вилочковой железы и открывается в правое предсердие. У взрослого скота она располагается в средостении, под плечеголовным стволом. Дополнительно в вену открывается рёберно-шейная вена, образующаяся слиянием глубокой шейной, позвоночной, дорсальной лопаточной и передней межреберной венами. В соустье их, в зависимости от диаметра, находятся, как правило, двух или трёхстворчатые клапаны.
С возрастом плодов и животных после рождения диаметр краниальной полой вены увеличивается неравномерно. Наиболее активно калибр вены возрастает в пять, восемь месяцев плодного и в шесть, и 12 месяцев постнатального периода, но наиболее интенсивно в возрасте три – семь дней и шесть месяцев после рождения. У крупного рогатого скота диаметр краниальной полой вены достоверно увеличи-вается до 18-месячного возраста, затем до трёх лет темп роста замедляется.
У крупного рогатого скота абсолютный прирост диаметра вены в первой половине плодного периода развития был значительно ниже, чем во второй.
Если рассматривать прирост по возрастным группам плодов, то выявляется следующая закономерность: у плодов он неравномерно возрастает до восьми месяцев и к рождению понижается (табл. 26).
26. Рост диаметра краниальной полой вены
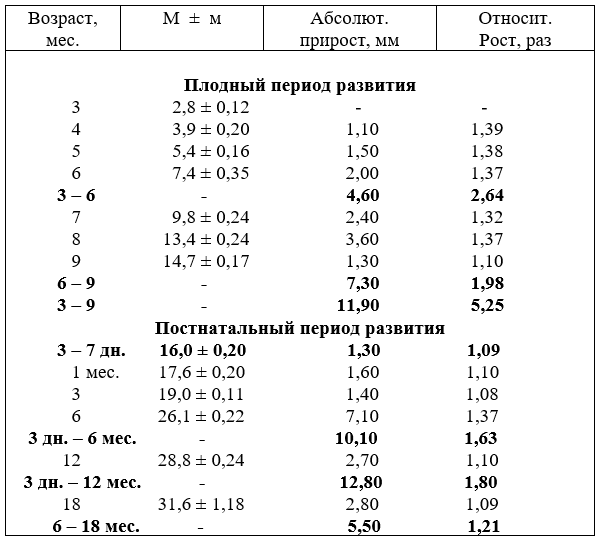
Несколько иная закономерность абсолютного прироста диаметра вены в постнатальном периоде развития – резко диаметр вены увеличивается в новорожденном возрасте и у телят в шесть месяцев, а затем темп прироста постепенно снижается. В целом в первом полугодии жизни телят прирост калибра вены был в два раза выше, чем в последующие 12 месяцев. Из данных видно как на рост вен влияет смена среды обитания, рода пищи и динамическая активность опорно-двигательного аппарата животных.
В относительных величинах наблюдается обратная зависимость роста диаметра краниальной полой вены по отношению к абсолютному приросту. Так,
в первую половину плодного периода развития относительный рост диаметра вены выше, чем во вторую половину и, соответственно, составляет 2,64 и 1,98
раза. По возрастным группам плодов относительный рост калибра вены был более равномерный, лишь к рождению несколько понижается. После рождения
относительный рост равномерно повышается до возраста шести месяцев и в этом возрасте молодняка достигает максимума. В следующие 12 месяцев рост калибра увеличивается только в 1,21 раза. Таким образом, в относительных
величинах темп роста диаметра вены, начиная с первой половины плодного периода развития и до 18 месяцев после рождения, плавно понижается и устанавливается в три года.
Вопросы для контроля
- Из слияния каких вен формируется плечеголовная вена?
- Из каких частей тела выносит кровь и куда открывается краниальная полая
вена?
- Перечислите вены, входящие в систему краниальной полой вены?
- Где располагается и откуда выносит кровь наружная ярёмная вена?
- На уровне каких шейных позвонков безопасно брать кровь из наружной
ярёмной вены?
- Как увеличивается диаметр наружной ярёмной вены и почему?
- Особенности расположения внутренней ярёмной вены?
- Какой вены следует считать продолжение краниальной полой вены?
- В каком возрасте устанавливается рост диаметра вен системы передней полой
вены?
9. СИСТЕМА КАУДАЛЬНОЙ ПОЛОЙ ВЕНЫ
Задняя полая вена отводит кровь в правое предсердие сердца из внутренних органов брюшной, тазовой полостей и их стенок, тазовых конечностей и половых органов. Образована слиянием общих подвздошных вен, в дорсальную стенку корня каудальной полой вены открывается средняя крестцовая вена. Располагается она вначале в брыжейке кишечника справа и несколько ниже от брюшной аорты, затем опускается к тупому краю печени. В вырезке печени принимает в себя печёночные вены, проходит через диафрагму в грудную полость ниже и справа от пищевода, проходит в средостении и открывается в венозный синус правого предсердия.
Из этого следует, что краниальная и каудальная полые вены несут потоки крови к сердцу друг другу навстречу, но гемодинамического столкновения их в правом предсердии не происходит, потоки крови разводит межвенозный валик (бугорок). Направляя поток крови, из краниальной полой вены в полость ушка, а из каудальной – в полость венозного синуса предсердия.
В полостях правого предсердия скорость потоков крови и давление резко падают, что способствует сохранению целостности клеток крови. По ходу в заднюю полую вену открываются глубокая окружная подвздошная, почечная,
надпочечниковые, в вырезке печени – печёночные, краниальная брюшная и каудальная диафрагмальная вены.
9.1. РОСТ ДИАМЕТРА ВНУТРЕННЕЙ ПОДВЗДОШНОЙ ВЕНЫ
Внутренние подвздошные вены парные и являются продолжением вперёд внутренних срамных вен. Они выносят кровь к сердцу из органов тазовой полости. Диаметр вен с возрастом плодов и животных после рождения увеличивается неравномерно и к рождению достигают 5,35 мм, а в 18 месяцев после рождения 10,12 мм (табл. 27).
Абсолютный прирост диаметра внутренних подвздошных вен с возрастом плодов тоже возрастает неравномерно. Активно он увеличивается в возрасте плодов четыре и семь месяцев, но к рождению несколько снижается. В шесть месяцев натального развития абсолютный прирост диаметра достигает 1,97 мм, а.
27. Рост диаметра внутренней подвздошной вены
в девять - 2,55 мм, т.е. в первую половину плодного развития значительно ниже, чем во вторую.
Несколько иные данные показывают рост диаметра вены относительные величины. Наиболее активно растёт диаметр вены в возрасте плодов четыре, пять
и семь, восемь месяцев, к рождению значительно снижается. Но в первую поло-
вину плодного развития диаметр вены возрастает в 3,37, а во вторую – только в 1,91 раза, что значительно ниже.
9.2. Рост диаметра наружной подвздошной вены
Наружные подвздошные вены парные и являются продолжением бедренных вен, выносят кровь из тазовых конечностей, брюшных стенок, вымени и половых органов. Диаметр вен с возрастом плодов и животных после рождения
увеличивается неравномерно и к рождению достигает 9,8 мм, а в 18 месяцев после рождения – 16,3 мм (табл.28).
28. Рост диаметра наружной подвздошной вены
Абсолютный прирост диаметра наружной подвздошной вены с возрастом плодов увеличивается неодинаково. Наиболее активно возрастает у плодов в
четыре, шесть и девять месяцев. В первую половину (3 – 6) плодного развития абсолютный прирост диаметра вены составил 3,54 мм, а во вторую – 3,80 мм, что выше, чем в первую.
Несколько другие данные показывают коэффициенты относительного роста диаметра вен. Так, наиболее активно диаметр вены увеличивается у плодов в четыре, шесть, семь и девять месяцев. Однако в первую половину плодного развития диаметр вен увеличивается в 2,44, а во вторую только – в 1,63 раза, что значительно меньше.
9.3. Рост диаметра общей подвздошной вены
Вена выходит из слияния внутренних и наружных подвздошных вен. Это
ствол у взрослых животных, длиной до 18 см и диаметром более 20 мм, поднимается по медиальной поверхности тела и крыла подвздошной кости косо
и вверх, и на границе V–VI поясничного позвонка правая и левая вены сливаются. Из их слияния вперёд справа и ниже от аорты выходит каудальная полая вена.
Диаметр общей подвздошной вены достоверно, но неравномерно увеличивается как в натальном, так и в постнатальном периодах развития и устанавливается в 18 месяцев (табл. 29).
29. Рост диаметра общей подвздошной вены
Наиболее активно диаметр ствола общих подвздошных вен возрастает к рождению и после рождения у телят до одного месяца. Такой рост диаметра общей подвздошной вены объясняется сменой среды обитания и мышечной нагрузкой направленной на поиск пищи, что стимулирует развитие диаметра вены особенно до шести месяцев после рождения. Затем калибр вены возрастает, но незначительно.
Как в плодном, так и в постнатальном периодах развития до возраста телят шести месяцев абсолютный прирост диаметра общих подвздошных вен изменяется неравномерно. Активно он возрастает в возрасте плодов четыре, шесть, семь и девять месяцев, особенно, в новорожденном и у телят в возрасте
одного месяца (табл.29), затем понижается. Но таких резких колебаний в приросте, которые описаны в системе краниальной полой вены нет. Как обычно максимальных величин в приросте диаметр достигает во второй половине плодного развития (4,85 мм) и значительно ниже в первой, но у телят после рождения до шести месяцев он увеличивается активнее, чем калибр внутренней и наружной подвздошных вен. Полагаем, что кроме приведённых выше причин, на их развитие оказывает половое созревание животных.
В относительных величинах рост диаметра общей подвздошной вены происходит более равномерно, чем в абсолютных величинах, но активнее возрастает в первой половине плодного развития, менее - во вторую и ещё слабее у телят до шести месяцев после рождения. Наиболее активно как в плодном, так и в постнатальном периодах развития увеличивает диаметр внутренняя подвздошная вена. Калибр наружной подвздошной и общей подвздошной вен растет почти одинаково. Эти обе вены связаны с кровоснабжением половых органов.
9.4. Рост диаметра воротной вены
Воротная вена выносит кровь из желудка, селезёнки, тонкого и толстого отделов пищеварительной системы. Исключение составляет прямая кишка, из которой кровь по внутренним подвздошным венам непосредственно поступает в каудальную полую вену, минуя печень. На этом обоснован метод введения лекарственных форм в организм через прямую кишку – per rectum.
Селезёночная, краниальная и каудальная брыжеечные вены, сливаясь, образуют короткий ствол под названием воротная вена, длиной до 10 см и диаметром до 28,3 мм, проходящий через ворота в печень. Отсюда её название «воротная вена». В печении она разветвляется на долевые вены, затем – междольковые, последние переходят во внутридольковые синусоидные капилляры печёночных долек. Капилляры внутри дольки образуют центральную вену дольки.
Центральные вены дольки открываются в поддольковые или собирательные вены, которые образуют долевые печёночные вены в количестве четырёх – шести. Они выходят из печени и в области вырезки для каудальной полой вены
открываются в заднюю полую вену. Из данного анализа следует, что печень стоит на пути оттока крови из пищеварительной системы. Поэтому, попавшие в кровь воротной вены токсические вещества, микроорганизмы из желудочного
кишечного тракта клетками печени нейтрализуются. В случае прорыва инфекции развивается тяжёлое заболевание печени.
Во всех возрастных группах плодного и постнатального периодов развития диаметр воротной вены увеличивается достоверно. В первой половине плодного развития калибр вены активно увеличивается в возрасте три и четыре месяца, затем понижается, во второй – резко возрастает в восемь и девять месяцев и максимума достигает концу беременности (табл. 30).
30. Рост диаметра воротной вены
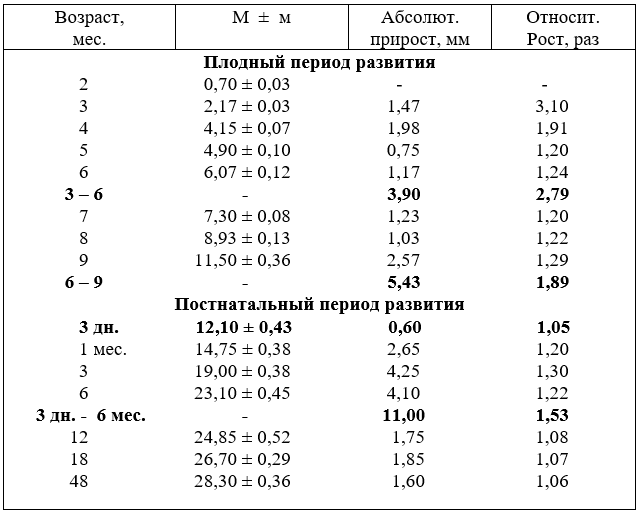
После рождения активный абсолютный рост диаметра вены продолжается до шести месяцев и достигает 23,1,0 мм. Во второй половине года темп роста замедляется, но продолжает увеличиваться до 18 месяцев.
Абсолютный прирост диаметра вены происходит волнообразно, но пики максимума отмечаются в три, четыре, шесть, семь и в девять месяцев плодного, и в три, шесть месяцев постнатального развития. Особенно бурно диаметр ворот-
ной вены увеличивается до возраста телят шести месяцев. Затем темп прироста диаметра падает и устанавливается в 18 месяцев. Но во вторую половину плодного развития был выше (5,43 мм), чем за первую (3,9 мм). Особенно резко возрастает диаметр воротной вены за первые шесть месяцев после рождения телят (на 11 мм), а до четырёх лет - только на 5,1 мм,
Относительные величины показывают, что в первую половину плодного развития диаметр воротной вены растёт более активно, чем во вторую половину. После рождения он активно продолжает увеличиваться до шести месяцев, затем темп роста плавно снижается. Эти морфометрические данные показывают как влияет на развитие плода, новорожденного, молодняка смена среды обитания, рода пищи, повышенная функциональная нагрузка на органы пищеварения и опорного двигательного аппарата.
9.5. Особенности роста диаметра каудальной полой вены
Задняя полая вена проходит через вырезку тупого края печени, срастаясь с ней, принимает кровь из печени. Затем проходит через одноименное отверстие в диафрагме и проникает в грудную полость. В полости она простирается к правому предсердию сердца в средостении, снизу и несколько справа от пищевода.
Диаметр задней полой вены у плодов и животных после рождения увеличивается неодинаково. Наиболее активно он возрастает к концу плодного периода развития и у молодняка до шести месяцев после рождения (табл. 31).
31. Рост диаметра (мм) каудальной полой вены
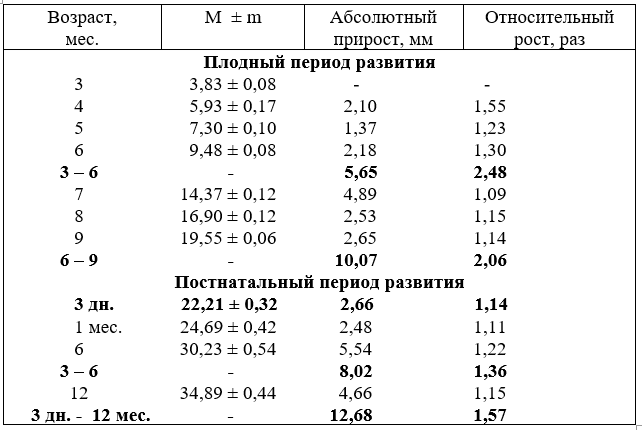
Абсолютный прирост диаметра в первую половину плодного развития увеличивается на 5,65 мм, во вторую – 10,07 мм, а за весь плодный период – 15,72 мм. За первые шесть месяцев постнатального развития прирост составил 8,02 мм, а за первый год жизни – 12,68 мм, что значительно ниже, чем за девять месяцев плодного периода развития.
В относительных величинах диаметр каудальной полой вены с возрастом плодов и животных после рождения увеличивается неравномерно, наиболее активно возрастает в четыре, шесть и восемь месяцев плодного развития и в
новорожденном, и у молодняка до шести месяцев - после рождения, затем плавно снижается.
При рассмотрении роста диаметра задней полой вены по этапам плодного и постнатального периодов развития, установлена следующая закономерность. В первой половине натального развития он увеличивается в 2,48, во второй – 2,06, а завесь плодный период развития - в 5,1 раза, после рождения за первое полугодие жизни телят в 1,36, за второе – 1,15, в за 12 месяцев – в 1,57 раза. Из
данного анализа следует, что наиболее активно диаметр задней полой вены возрастает в первую половину плодного развития и менее – во вторую, за весь плодный период в 5,1 раза, что значительно больше, чем за первое и второе полугодие жизни молодняка после рождения.
Из сравнения роста диаметров краниальной (табл.26) и каудальной (табл.31) полых вен видно, что диаметр их увеличивается неодинаково. В плодном развитии, в абсолютных величинах, активно увеличивает диаметр задняя, а после
рождения - передняя полая вена, в относительных величинах – во всех периодах передняя полая вена развивается активнее, исключение составляет первая половина плодного развития и шесть месяцев после рождения.
9.6. Морфофункциональное обоснование роста полых вен
В абсолютных величинах прирост диаметра полых вен активно возрастает во вторую половину плодного периода развития и слабее – в первую (табл.32). Но
задней полой вены прирост диаметра выше в любой возрастной группе плодов, чем передней.
После рождения, до возраста телят шести месяцев диаметр передней полой вены увеличивается уже активнее, чем задней. Диспропорция в росте калибра
вен постепенно выравнивается к возрасту молодняка 12 и устанавливается в 18 месяцев.
32. Абсолютный прирост и относительный рост полых вен
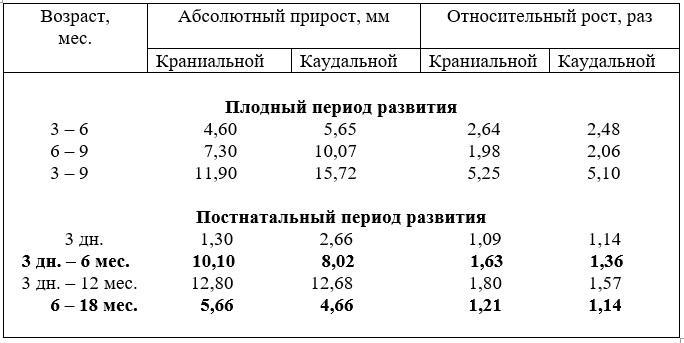
В относительных величинах рост диаметра полых вен активно возрастает в первую половину плодного развития и краниальной полой вены выше, чем каудальной. В целом калибр вен активно увеличивается в плодном развитии, чем после рождения.
В постнатальном периоде развития диаметр вен интенсивно растёт у телят в возрасте шесть и 12, и устанавливается в 18 месяцев и всё же темп роста диаметра краниальной полой вены выше, чем каудальной. По видимому на рост передней полой вены влияет развитие головного мозга, мышц передней части тела и желез внутренней секреции, что требует дополнительных исследований.
Вопросы для контроля
- Объединение каких вен формирует каудальную полую вену?
- Из каких частей тела выносит кровь каудальная полая вена?
- Особенности топографии задней полой вены?
- Какие вены открываются в брюшной полости в заднюю полую вену?
- Особенности формирования и роста воротной вены печени?
- Куда приносит кровь задняя полая вена?
- Откуда выносят кровь внутренние подвздошные вены?
- Что такое общие подвздошные вены и что они формируют?
- Особенности роста диаметра вен, составляющих систему задней полой вены?
- Особенности формирования и роста воротной вены печени?
- На чём обоснован метод введения лекарственных средств - per rectum?
- Особенности роста диаметра каудальной полой вены?
- Дайте обоснование роста задней и передней полых вен?
- Из чего возникла дуга аорты в филогенезе?
- Из чего возник артериальный проток в филогенезе?
- Во что превращается артериальный проток в онтогенезе?
- Во что в филогенезе превратилась шестая жаберная артерия?
- Какой сегмент дуги аорты способствует опережающему росту передней части тела?
- Когда наиболее активно увеличивается диаметр артерий?
- Как развивается грудная аорта относительно брюшной аорты?
- Как развиваются артерии передней и задней частей тела относительно друг друга?
- Почему задняя часть тела отстаёт в развитии от передней части и, что этому способствует?
- Как растут диаметры полых вен относительно друг друга?
- В каком возрасте выравнивается диспропорция в росте полых вен?
- Сердечнососудистая система возникает рано из мезенхимы и её производных, и развивается со значительным опережением по сравнению с развитием других органов, т.к. она доставляет для их развития питательные вещества.
- В зародышевом, предплодном и раннем плодном периодах развития создаётся жидкостная среда и «поле роста» для дифференциации клеток, закладки и формирования органов. Создание таких условий в формирующемся организме возможно тогда, когда приток крови по артериям будет превалировать над её оттоком по венам – это закон. По этому, в эти периоды активно растёт диаметр артерий и слабо вен. В связи с этим, предполагаем, что рост организма может продолжаться до тех пор, пока натяжение тканей не выровняется с давлением крови. При этом генную теорию мы не отрицаем.
- Во второй половине плодного периода развития органы и системы плода уже сформированы и функционируют, вследствие чего, в них накапливаются продукты метаболизма, которые можно вывести к выделительным органам и нейтрализовать их путём интенсивного оттока крови по венам, что способствует усиленному росту диаметра вен.
- В предродовом периоде развития идёт подготовка организма к смене среды обитания, масса плода за месяц до рождения удваивается, что требует больших затрат питательных веществ, приносимых током крови. В связи, с чем у поздних плодов активно растёт диаметр артерий.
- В новорожденном периоде развития произошла смена благоприятной, внутриутробной среды обитания, на агрессивную воздушную среду, по этому
- Как растут артерии относительно вен в онтогенезе?
- К какому возрасту выравнивается диспропорция в росте артерий вен?
- Как развиваются полости желудочков сердца относительно друг друга?
- Какие причины вызывают подобный рост кровеносных сосудов?
- Как меняется структура компактного костного вещества костей конечностей плодов и телят после рождения?
- Зависит или нет от функциональной нагрузки структура компактного костного вещества периферического скелета?
- Как действую гиперфункция и гипофункция на структуру костей?
- Как изменяется толщина суставных хрящей под действием гиперфункции?
- Как влияет суставный угол на толщину хрящей проксимальных и дистальных эпифизов трубчатых костей конечностей?
10. Морфофункциональное обоснование возрастных особенностей артерий передней части тела
Как видно из главы 2,1, диаметр сегментов дуги аорты увеличивается неодинаково как в целом составляющих дугу, так и в отдельности. Особенно значительным колебаниям в росте подвержен второй сегмент относительно артериального протока и третьего сегмента дуги аорты (см.табл.3). Чем объяснить подобный рост? На основании собственных исследований можно утверждать, что артериальный проток (связку) в филогенезе следует рассматривать как сохранившийся отрезок (остаток) бывшей спинной аорты между V и VI, а второй сегмент – между IV–V жаберными артериями. Из этого следует, что дуга аорты в целом возникла из отрезков спинной аорты между IV
и V межжаберными артериями, а артериальный проток, который к 1,5 месячному возрасту в норме запустевает и превращается в связку, является остатком спинной аорты между V и VI жаберными артериями. VI-я жаберная артерия превратилась в легочной ствол, из которого впоследствии возникли правая и левая легочные артерии.
Из этого следует, что артериальный проток плодов относительно третьего сегмента аорты, выполняет большую динамическую нагрузку, чем второй сегмент. Узкий просвет, которого, в плодном периоде развития способствует местному сопротивлению движения крови из первого сегмента в третий, иначе
из левого желудочка сердца в грудную аорту. Сформированная «горловина» на месте второго сегмента, способствует интенсивному кровотоку из луковицы аорты в плечеголовной ствол плода. Одновременно поток крови, идущий из артериального протока в дугу аорты, способствует оттоку крови из первого сегмента в третий через «горловину» по принципу инжектора. Отсюда следует, что второй сегмент в плодном периоде развития выполняет роль анастомоза между первым и третьим сегментом, возникший в филогенезе из участка дорсальной аорты между IV-V жаберными артериями. Лишь после облитера-
ции артериального протока он выполняет в полном объёме проведение крови из левого желудочка сердца в грудную аорту.
Диаметр плечеголовного ствола с возрастом плодов и животных после рождения увеличивается более равномерно, чем калибр сегментов дуги аорты и стабилизируется в возрасте 18 месяцев после рождения. Но внешний и внутренний диаметры плечеголовного ствола увеличиваются неодинаково, что связано с неравномерностью роста толщины стенок и у быков он развивается
интенсивнее, чем у тёлок. Стабилизация толщины стенок их у быков идёт активнее, чем у тёлок (табл.5,6) и устанавливается относительно друг друга в возрасте 18 месяцев.
Диаметр подключичных артерий самым широким, по отношению к плечеголовному стволу, был у плодов трёх месяцев (62,2%). Затем развитие диаметра к рождению и после рождения понижается и у телят в шесть месяцев составляет 39,9%, К возрасту 18 месяцев он увеличивается до 54,7%. Считаем,
это будет показано ниже, что телята в молозивном, молочном, переходном периодах развития недостаточно подвижны. С переходом на грубые корма динамическая нагрузка на мышечную систему повышается (ноги кормят животного), что сказывается на развитии подключичной артерии (табл.7).
Диаметр общего ствола сонных артерий с возрастом неодинаково превосходит калибр подключичных артерий (табл.8). Так, он в плодном периоде развития превышает диаметр подключичных артерий в 1,49 и в постнатальном –
в 1,34 раза и относительно плечеголовного ствола остаётся самым широким в возрасте плода в три месяца (95,7%). К рождению понижается до 73,7% и на этом уровне остаётся у телят до трёх месяцев после рождения, затем волнообразно повышаясь и понижаясь, стабилизируется в 18 месяцев (79,8%)
. Из данного анализа следует, что ствол развивается с опережением по отношению к подключичным артериям. Полагаем, что на его рост влияет развитие головного мозга, черепа и мышц шеи, головы. Поэтому во всех возрастных группах плодного и постнатального периодов развития продолжением плечеголовного ствола следует считать общий ствол сонных артерий.
Диаметр общих сонных артерий неравномерно увеличивается до 18 месяцев. Особенно активно он возрастает у новорожденных телят. У быков диаметр увеличивается интенсивнее, чем у тёлок и разница в диспропорции их роста выравнивается к 12 месяцам после рождения. Диаметр остальных артерий шеи неравномерно увеличивается до 18 месяцев постнатального развития. Особенно активный рост калибра их артерий наблюдается у новорожденных телят.
В среднем в первую половину плодного периода развития абсолютный прирост диаметра артерий передней части тела достигает 3,80, во вторую – 4,27 мм, а после рождения до шести месяцев – 4,3 мм и устанавливается в 18 месяцев (табл.33).
33. Абсолютный прирост диаметра (мм) артерий передней части тела
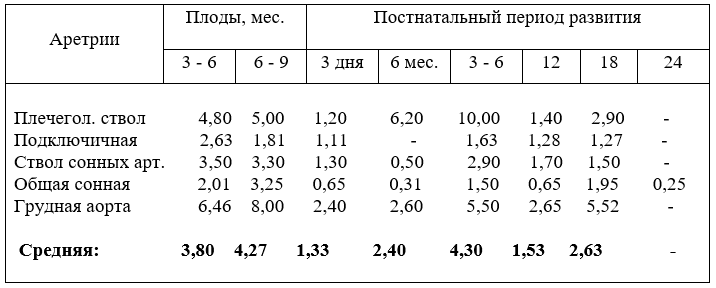
Из таблицы видно, что наиболее активно в среднем диаметр артерий увеличивается во вторую половину плодного периода развития (4,27 мм) и до
шести месяцев у молодняка после рождения (4,3 мм), устанавливается прирост диаметра в 18 месяцев. Затем увеличивается, но незначительно.
В научной литературе имеются работы по морфофункциональному развитию сердечнососудистой системы (И.В.Хрусталёва, 1972; Б.П.Шевченко, 1999; Х.Б.Баймишев, 2001; М.С.Сеитов, 2001; М.Ж. Нурушев, Б.П. Шевченко и др., 2010). Все они свидетельствуют о неравномерности прироста их как в натальном, так и в постнатальном периодах развития. Однако фактический материал, характеризующий рост кровеносной системы в возрастном аспекте беден и не позволяет сделать какие либо количественные обобщения. Полученные экспериментальные данные позволяют нам компенсировать этот недостаток.
Если рассматривать рост диаметра магистральных артерий в относительных величинах, то прослеживается обратная зависимость по отношению к абсолют-
ному приросту. Так, диаметр артерий передней части тела интенсивнее развивается в первую и значительно ниже во вторую половину плодного периода. Например, в первую половину плодного развития диаметр артерий возрастает в 5,03, а во вторую – 2,57 раза (табл. 34).
34. Относительный рост диаметра (раз) артерий передней части тела
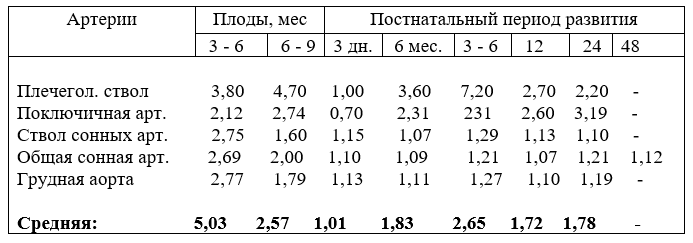
После рождения калибр артерий интенсивно возрастает у молодняка до шести месяцев. За первые полгода жизни калибр артерий увеличивается в 2,65
раза, а за второе полугодие только в 1,72 раза. Относительный рост сосудов устанавливается в два года после рождения. Затем рост калибра артерий увеличивается до четырёх лет, но незначительно.
Все артерии краниальной части тела имеют магистральный тип ветвления и образуют внутрисистемные и межсистемные анастомозы и коллатерали. Внутрисистемные анастомозы поверхностной и глубокой шейной артерии в фасциях, в фасциальных влагалищах и наружном перимизии мышц образуют грубые, крупно-петлистые сети, внутри мышц – тонкие, мелко-петлистые, ячеи которых вытянуты по ходу мышечных пучков.
Вопросы для контроля
11. Морфофункциональное обоснование возрастных особенностей артерий задней части тела
Последовательность хода ветвления артерий брюшной, тазовой полостей
исследовали Н.А.Малков (1979), Р.Ш.Тайгузин (1985), М.В.Андреев (1988). Установлено, что энергия роста сосудов в онтогенезе происходит неравномерно. В плодном периоде развития наиболее активно диаметр сосудов возрастает в четыре, семь и девять месяцев и к рождению достигает максимальных величин, в постнатальном - в три дня и шесть месяцев.
Морфометрические показатели роста артерий задней чести тела крупного
рогатого скота показывают, что диаметр грудной, брюшной аорты и их ответвлений с возрастом плодов и животных после рождения увеличивается неравномерно (табл.10–15) и устанавливается в 18 месяцев. Затем возрастает, но незначительно.
Абсолютный прирост диаметра грудной, брюшной аорты, как и их дериваций, максимальных величин достигает во второй и значительно ниже – в первой половинах плодного периода развития. Но абсолютный прирост диаметра грудной аорты значительно выше в первой (6,46) и второй (8,0 мм) половинах плодного развития по сравнению с брюшной аортой, соответственно – 5,63 и 5,66
мм. Из данного анализа следует, что диаметр брюшной аорты в развитии запаздывает по отношению к грудной аорте. Считаем, что это связано с задержкой развития в первой половине плодного периода пищеварительной, мочевыделительной систем и в целом задней части тела (табл.35).
35. Абсолютный прирост диаметра (мм) артерий задней части тела
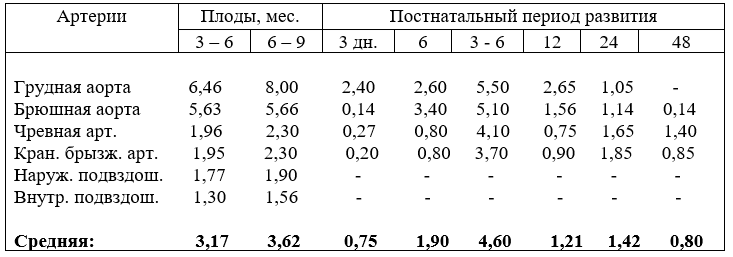
В постнатальном периоде развития диаметр грудной аорты продолжает активнее расти, чем брюшной, и только с месячного возраста телят рост диаме-
тра брюшной аорты усиливается, и разница в диспропорции их выравнивается в шесть месяцев. Однако диаметр грудной аорты продолжает активнее развиваться и до 18 месяцев, а брюшной – как бы запаздывает в приросте и выравнивается только к двум годам.
После рождения активно возрастает диаметр всех артерий телят до шести месяцев. Но абсолютный прирост в первом полугодии после рождения был значительно выше, чем с шести до 12 месяцев. Появляется предположение, что кровеносная система спешит заложить запас объёма сосудов на случай повышения динамических нагрузок с возрастом на органы, особенно, на будущий зимний сезон. Причём абсолютный прирост диаметра грудной аорты значительно превышает прирост диаметра брюшной аорты. Исключение составляют телята в возрасте шести месяцев.
В относительных величинах просматривается обратная зависимость роста артерий задней части тела по отношению к абсолютному приросту. Считаем, что абсолютный прирост не всегда полно раскрывает сущность функционального становления сосудов, а более точно раскрывают относительные, независимо в каких величинах они выражены - разах, процентах и т.д. Разница заключается в методике их обсчёта.
Относительные величины показывают, что у плодов до шестимесячного возраста активнее развивается брюшная аорта, а во второй половине плодного развития и у телят после рождения до трёх дней – грудная аорта, затем до шести месяцев – вновь брюшная аорта (табл.36).
36. Относительный рост диаметра (раз) артерий задней части тела
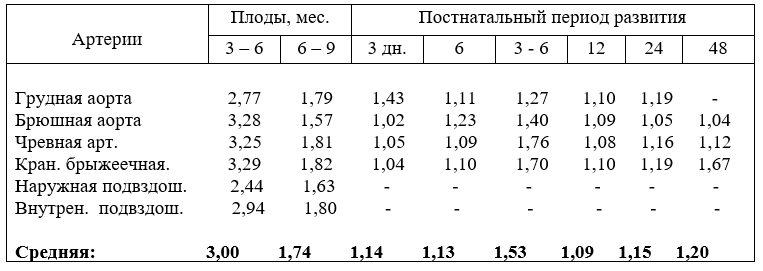
В целом артерии задней части тела крупного рогатого скота интенсивнее развиваются в первую половину плодного периода развития, чем во вторую, что способствует отставанию в росте задней части тела от передней. Однако после рождения до шести месяцев их рост выравнивается. Из данного анализа следует, что артериальные сосуды передней и задней частей тела, в целом, развиваются одинаково, исключение составляет брюшная аорта, развитие которой попадает под влияние морфофункционального развития органов брюшной и тазовой полостей, поэтому несколько запаздывает в росте.
Вопросы для контроля
12. Морфофункциональное обоснование возрастных особенностей вен передней части тела
В краниальную часть тела кровь поступает из плечеголовного ствола, а отводится по краниальной полой вене в правое предсердие сердца. Из головы
выносит кровь наружная ярёмная и в 37% случаев ей помогает внутренняя ярёмная вена, которые сливаясь, формируют общий ствол ярёмных вен.
Из шеи кровь выносят позвоночная, глубокая шейная, дорсальная лопаточная и передняя межреберная вены, Объединяясь, они образуют реберную шейную вену, которая открывается в дорсолатеральную стенку краниальной полой вены. В соустье их находится трёхстворчатый клапан.
Подключичная вена, диаметром 1,75–17,61 мм, выносит кровь из грудных стенок и конечностей. Открывается она в боковую стенку плечеголовной вены на границе перехода в краниальную полую вену. В соустье их находится трёхстворчатый клапан. Диаметр вены достоверно увеличивается до 18 месяцев
после рождения. Хотя рост диаметра её в 18 месяцев достоверен, но он значительно уступает скорости роста до года (см. табл.25).
У крупного рогатого скота абсолютный прирост диаметра подключичной вены в первой половине плодного периода развития был значительно ниже, чем во второй. После рождения диаметр вены активно возрастает у телят до возраста
шести месяцев, но значительно ниже, чем у плодов непосредственно перед рождением. Диаметр стабилизируется в росте у молодняка 18 месяцев. Затем до трёх лет возрастает, но незначительно (табл.37).
37. Абсолютный прирост диаметра (мм) вен передней части тела
Прирост диаметра наружной ярёмной вены, в целом, идентичен подключичной вене, исключение составляет возраст шесть месяцев. Абсолютный прирост диаметра позвоночной и глубокой шейной вен увеличивается одинаково, максимальный прирост в плодном развитии, приходится на конец второй половины и на шесть месяцев постнатального периода. Диаметр обеих вен устанавливается в 18 месяцев после рождения.
Наиболее интенсивно, по сравнению с другими венами, возрастает в абсолютных величинах диаметр краниальной полой вены. В первой половине плодного развития диаметр вены прирастает на 4,6 мм, к концу второй половины
– на 7,25 мм, а за шесть месяцев после рождения – на 11,47 мм. Ни один сосуд в организме не имеет таких высоких темпов прироста как краниальная полая вена.
Затем темп абсолютного прироста понижается и устанавливается в два года.
И так, в среднем абсолютный прирост диаметра вен выдерживает закономерность свойственную артериям. Разница заключается в том, что прирост диаметра вен, начиная с ранних плодов, постоянно возрастает до шести месяцев после рождения, а затем понижается и устанавливается в два года. У артерий, наоборот, начиная с ранних плодов и до шести месяцев после рождения, понижается и устанавливается в 18 месяцев.
В относительных величинах наблюдается обратная закономерность роста диаметра вен по отношению к абсолютному приросту, свойственному артериям.
Так, в первую половину плодного развития относительный рост диаметра вен выше, чем во вторую и, соответственно, составляет 2,22 и 2,04 раза. Особенно высокий рост в эту половину происходит краниальной полой вены (2,64), подключичной (2,53) и наружной ярёмной вены (2,34 раза).
38. Относительный рост диаметра (раз) вен передней части тела
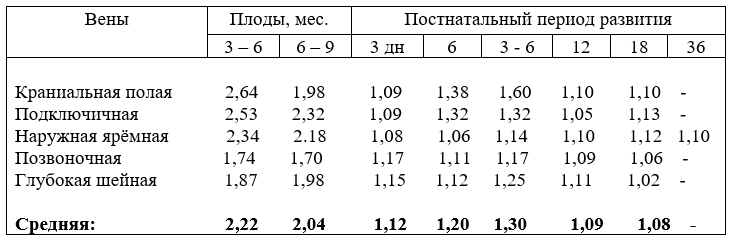
В постнатальном периоде развития высокий темп относительного роста диаметра вен сохраняется у молодняка до шести месяцев. В целом за первое полугодие жизни телят темп роста диаметра вен был выше, чем во втором (до 12 месяцев, табл.38).
Вопросы для контроля
1. Откуда выносит кровь подключичная вена и куда?
2. В каком возрасте устанавливается рост диаметра вен системы передней полой вены и артерий плечеголовного ствола?
13. Морфофункциональное обоснование возрастных особенностей вен задней части тела
В заднюю часть тела кровь поступает по аорте, а возвращается к сердцу по каудальной полой вене. У крупного рогатого скота задняя полая вена выходит из слияния правой и левой общих подвздошных и средних крестцовых вен. Располагается она в брыжейке кишки несколько слева и ниже от аорты, а при наполненном рубце может сдвигаться вправо. По ходу вена принимает приточные: почечные, надпочечниковые, краниальную брюшную, каудальную диафрагмальную вены. При прохождении через тупой край печени в неё открываются печеночные вены. Затем через отверстие в куполе диафрагмы проходит в средостение грудной полости и открывается в правое предсердие.
Абсолютный прирост диаметра магистральных вен задней части тела по возрастным группам плодов и животных после рождения прирастает неодинаково. Наиболее интенсивно он увеличивается во второй половине плодного и до шести месяцев постнатального периодов развития. В 18-месячном возрасте прирост диаметра всех вен понижается, но в старших возрастных группах может незначительно повышаться.
Наиболее активно диаметр вен прирастает в возрасте плодов от шести до девяти и у телят после рождения до шести месяцев. Если в плодном периоде развития воротная вена в росте диаметра отстаёт от задней полой и общей подвздошной вены, то после рождения до возраста телят шесть месяцев она резко увеличивает прирост и выходит на первое место (11,0 мм), а на второе - каудальная полая вена (8,02 мм).
39. Абсолютный прирост диаметра (мм) вен задней части тела
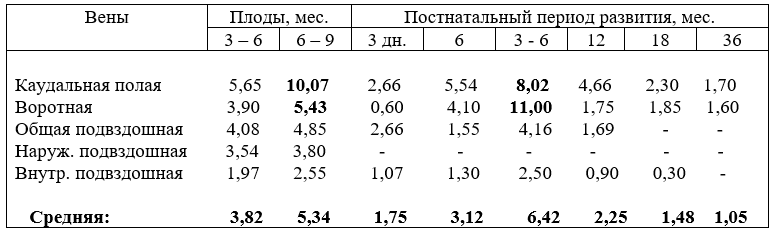
В целом абсолютный прирост диаметра вен остаётся высоким в возрасте плодов от шести до девяти месяцев (5,34) и у телят до шести месяцев после рождения (6,42 мм), что выше по сравнению с венами передней части тела. Но задняя полая вена активнее прирастает у плодов, а передняя – у телят до шести месяцев после рождения (табл.37,39).
В относительных величинах просматривается общая закономерность роста диаметра вен, свойственная венам передней части тела. Так, в первую половину плодного периода развития относительный рост диаметра вен выше, чем во вторую и, соответственно, достигает 2,70 и 1,87 раза. Но рост каудальной полой вены в относительных величинах ниже, чем краниальной полой и, соответственно, составляет 2,64 и 2,48 раза. Из системы вен задней части тела усиленно к концу беременности растёт диаметр каудальной полой вены и у телят до шести месяцев - воротной, затем, внутренней подвздошной вены. Относительный рост их устанавливается в 18 месяцев (табл.40).
В заключение следует отметить, что в абсолютных величинах диаметр полых вен активно возрастает во вторую половину плодного периода развития: краниальной – на 7,3 мм, а каудальной, соответственно – 10,07 мм (табл. 37,39).
40. Относительный рост диаметра (раз) вен задней части тела
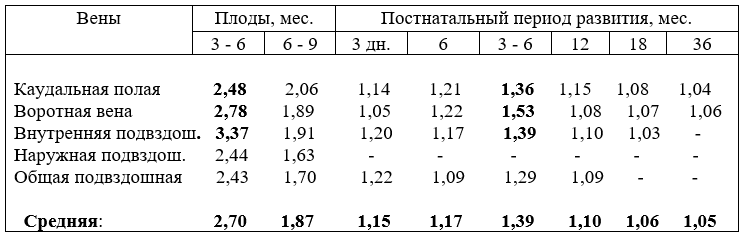
Всё же диаметр каудальной полой вены в этом развитии интенсивнее прирастает, чем краниальной полой вены, соответственно - 11,90 и 15,72 мм
(табл.41). После рождения до шести месяцев диаметр краниальной полой вены опять возрастает активнее, чем каудальной. Диспропорция в росте диаметра
полых вен постепенно выравнивается к возрасту животных 12 и устанавливается
в 18 месяцев.
41. Абсолютный прирост (мм) и относительный рост (раз) полых вен
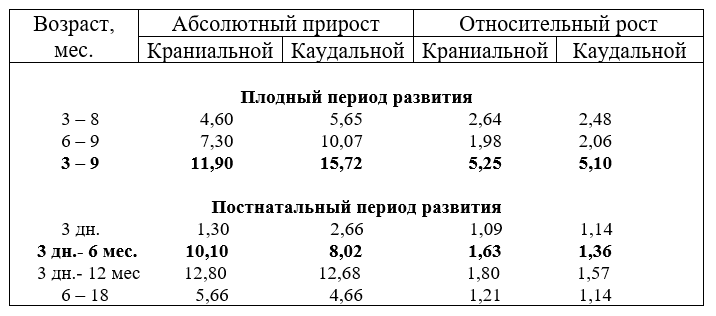
В относительных величинах рост диаметра полых вен, наоборот, активно возрастает в первую половину плодного развития и краниальной полой вены интенсивнее, чем каудальной, исключение составляют новорожденные. В целом, калибр вен активнее прирастает в плодном периоде развитии, чем в постнатальном (табл.41).
Вопросы для контроля
14. Рост диаметра артерий относительно вен
В специальной литературе имеются работы, раскрывающие развитие и морфофункциональное становление кровеносных сосудов в онтогенезе (Ф.И.Валькер, 1954; А.И.Никифоров, 1970). Однако этого недостаточно. Поэтому
фактический материал, характеризующий сосудистую систему в возрастном аспекте, беден и не позволяет решать какие-нибудь качественные и количественные обобщения, а тем более практические задачи. Полученные данные от магистральных сосудов тела крупного рогатого скота, северного оленя, белых крыс позволяют, в какой-то степени, компенсировать этот пробел белых страниц в биологической науке.
В онтогенезе животное постоянно испытывает влияние, с одной стороны, меняющейся внешней, а с другой - внутренней среды собственного организма, что видно на примере развития половой клетки, эмбриона, плода и, наконец, животного после рождения. Воздействие меняющейся внешней среды на развивающийся организм ещё изучено недостаточно, хотя исследования в данном направлении постоянно ведутся (А.П.Жуков, 2001).
Внутренняя среда организма зависит от рода пищи, воды и геопровинции, в которой обитает животное. В формировании её огромная роль принадлежит пищеварительной, мочевыделительной, сердечной сосудистой и лимфатической системам. Кровеносная и лимфатическая системы принимает непосредственное участие в создании внутренней среды. Ю.Т.Техвер (1972) пишет, что организм живёт в «текучей воде», под «текучей водой» он понимает, движущуюся кровь по сосудам. К этому следует добавить и «развивается» (Б.П.Шевченко, 1982), но вопрос формирования внутренней среды в онтогенезе ещё не изучен - это одна из актуальных задач, представляющая большой научный и практический интерес.
По росту диаметра артерий относительно диаметра вен имеется лишь несколько работ, выполненных на плодах человека (Н.В.Попова-Латкина, Е.П.Колоколова, Т.М.Щербакова, 1975), крупного рогатого скота и северного
оленя (Б.П.Шевченко, 1999). О том, что исследование в таком направлении сосудистой системы имеет не только теоретическое, но и большое практическое
значение указывали Г.М. Удовин (1970), М.Г. Привес (1974), И.В.Хрусталёва (2001). Но проблема роста объёма, длины, диаметра артерий к равнозначной вене, как способ оценки функционального состояния кровеносной системы, не решена до наших дней указывают А.И. Никоноров (1970), О.В. Алексеев (1981), Г.Н. Кассиль (1983), Б.П. Шевченко (1999), Х.Б. Баймишев (2001).
Особенно это хорошо видно на примере роста диаметров общей сонной артерии и наружной ярёмной вены (табл. 42).
42. Динамика роста диаметров общей сонной артерии и наружной ярёмной вены
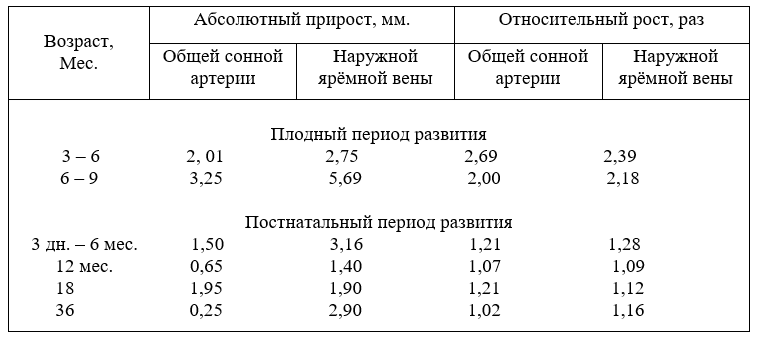
Два основных кровеносных сосуда – по первому кровь поступает от сердца к голове, по второму отводится от головы к сердцу. В абсолютных величинах порирост калибр артерии и вены во второй половине плодного развития выше (3,25 и 5,69 мм), чем в первой (2,01 и 2,75 мм) и наружной ярёмной вены диаметр значительно превосходит калибр общей сонной артерии и это так. Поэтому абсолютные величины свидетельствуют слабо о функции органа. Однако относительные величины показывают, что диаметр артерии и вены в первой половине плодного развития (3 – 6 мес.) развивается интенсивнее, чем во второй (6 – 9 мес.). Причём калибр общей сонной артерии в первой половине плодного развития значительно опережает рост диаметра (2,69 раза) наружной ярёмной вены (2,39 раза). Рост их диаметра относительно друг друга устанавливается в возрасте телят шесть месяцев после рождения. В старших возрастных группах с
переменным успехом увеличивается до 18 месяцев. С возраста животных 18 месяцев диаметр общей сонной артерии почти не возрастает (0,25 мм), а вот наружной ярёмной вены продолжает увеличиваться (2,9 мм).
Из сравнительных данных роста диаметра артерий и вен передней и задней частей тела видно, что диаметр вен в зрелом возрасте животных превосходит диаметр артерий в среднем в два (1,8–2,2) раза. Но с возрастом плодов и животных неодинаково. У плодов усиленно развиваются артерии передней части тела, а задней - отстают, но вены задней части, наоборот, опережают рост вен передней части тела (табл.43). Данное сравнение хорошо поясняет отставание в развитии задней части тела новорожденных телят.
43. Сравнительный абсолютный прирост диаметра (мм.) артерий и вен передней и задней частей тела
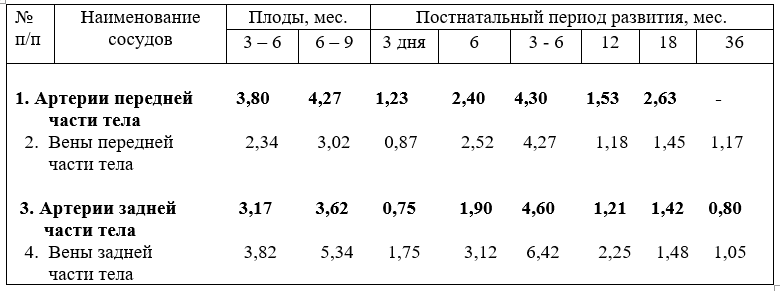
Р.Я. Козаченко (1987) установлено, что в предплодном и раннеплодном периодах развития крупного рогатого скота стенка аорты толще диаметра её просвета, а с повышением возраста внутриутробного развития эти соотношения становятся обратными. Выявлена закономерность опережающего роста аорты по отношению к каудальной полой вене, что создаёт благоприятные условия для развивающегося организма в утробе матери и создания «поля роста». Об этом
свидетельствуют не только абсолютные, но и показывают относительные величины. Артерии передней части тела в первую половину плодного периода развития увеличиваются в 5,03, во вторую – 2,57 раза, а вены, соответственно – в 2,22 и 2,04 раза. Эта диспропорция в росте выравнивается постепенно к возрасту 18 месяцев после рождения (табл.44).
44. Относительный рост диаметра (мм.) артерий и вен передней и задней частей тела
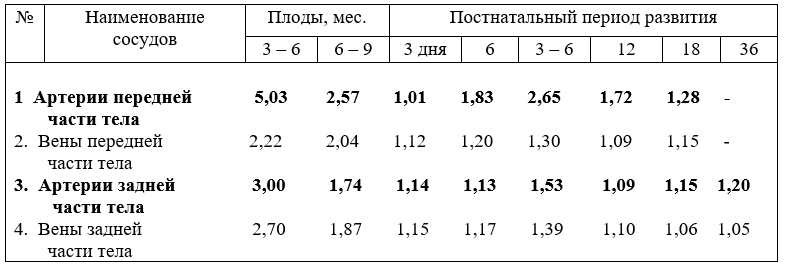
Артерии задней части тела плодов интенсивнее развиваются в первую половину плодного периода развития. Начиная со второй половины и до 12 месяцев после рождения, артерии и вены в относительных величинах растут одинаково. По этому, в отставании развития задней части тела играют роль не только и не столько провизорные образования (аранциев, артериальный протоки и овальное отверстие сердца и др.) сколько развитие кровеносных сосудов и полостей сердца (Я.Т. Подковыров, 1969). И так, из краткого анализа роста диаметра артерий относительно вен, или наоборот, следует, что с возрастом плодов и животных после рождения до года увеличивается волнообразно и устанавливается в 18 месяцев.
Наши данные по росту сосудов крупного рогатого скота, северного оленя и
крыс согласуются с развитием полостей правой и левой половин сердца, установленных профессором Я.Т. Подковыровым (1974) и доцентом Э.М.Бикчентаевым (1977). Авторы справедливо доказывают, что в раннем плодном периоде развития активно увеличивается масса и объём левой половины
сердца, а правая в это время отстаёт в развитии. В позднем плодном периоде развития, наоборот, рост правой половины сердца обгоняет левую, но к рождению масса и объём левой половины вновь опережает правую. Ясно какое значение имеет развитие объёма полостей сердца, т.к. из левой половины сердца выходит большой круг кровообращения, а из правой – малый.
С проблемами целостности организма и его интегрирующих систем, эволюционной морфологии и жизненных циклов можно познакомиться в трудах
Г.М.Удовина (1972), А.Н.Студитского (1981), И.И.Шмальгаузена (1982). По их данным, животный организм - это не «застывшая масса» клеток, тканей и органов, а постоянно развивающаяся и меняющаяся в онто-филогенезе сложная и взаимозависимая живая структура. Она может постоянно совершенствоваться по биологическим законам и под действием внешних, и внутренних факторов.
Организм строится на протяжении жизни как единое целое из морфологически связанных живых структур. Их природа такова, что они постоянно сами разрушаются, восстанавливаются и совершенствуются.
В онтогенезе животное испытывает влияние, с одной стороны, меняющейся внешней, а с другой – внутренней среды собственного организма, что видно на примере развития половых клеток, эмбриона, плода и, наконец, наворожденного телёнка, молодняка и т.д. Воздействие меняющейся внутренней и внешней среды на организм животных ещё недостаточно изучен.
Внутренняя среда организма зависит от рода пищи, воды и геопровинции, в которой животное обитает. В формировании её огромная роль принадлежит пищеварительной, мочевыделительной и, особенно, сердечной сосудистой системам. Кровеносная и лимфатическая система принимает непосредственное участие в создании внутренней среды, понятно, что организм живёт в этой «текучей воде» и «развивается», но вопрос формирования внутренней среды и
воздействия её на организм развивающегося животного в онтогенезе ещё слабо изучен – это одна из актуальных задач, представляющая большой научный и практический интерес, ждёт своего решения.
Таким образом, в первую половину плодного периода развития, когда артерии развиваются активнее вен, возникают условия замедленного возвращения крови к сердцу, усвоение питательных веществ, приносимых током крови, в это время
будет более благоприятным и полным. В то же время, токсические вещества, попавшие через плаценту в организм плода, будут действовать на молодые клетки формирующегося организма продолжительнее, чем могут вызвать неполноценное развитие отдельных органов, систем и организма в целом или смерть, что часто встречается в практике животноводства. Во второй половине
плодного периода развития, когда вены развиваются интенсивнее, происходит более быстрое по времени «вентилирование» крови в замкнутой сосудистой системе, способствующее нейтрализации и выведению токсических веществ из организма плода, что было доказано опытным путём (Б.П.Шевченко, 1982).
Волнообразный рост желудочков сердца и диаметра артерий, вен в плодном периоде развития относительно друг друга, можно объяснить следующими причинами:
идёт адаптация новорожденного к агрессивной внешней среде, что также требует значительных затрат питательных веществ. Последнее обстоятельство приводит к опережающему росту диаметра артерий относительно вен. Однако в половозрастном периоде диаметр вен больше артерий в 1,8–2,2 раза.
Кровеносная система представляет систему сообщающихся трубок, подразделяющуюся на артерии, микроциркуляторное русло и вены. Вся эта структура функционирует в тканях, органах и организме – как единое целое, и решать её проблемы следует комплексно по схеме: сосуд – орган – сосуд.
Полученные данные могут служить тестом научных обоснований при составлении рационов стельным коровам.
Прежде, чем перейти к рассмотрению кровоснабжения и васкуляризации костей конечностей, хотелось рассмотреть их структуру под действием различных факторов.
Вопросы для контроля
15. Структура костей конечностей в зависимости от двигательной активности
Развитие морфологии животных непосредственно связано с доместикацией животных. Задачи, которые выдвигает практика, являются основополагающими для неё - как фундаментальной науки, призванной устанавливать закономерности
роста, развития адаптационных возможностей структур организма на системном, макро–микро анатомическом уровнях и решать их в практическом аспекте.
Выяснение оптимальных условий содержания животных в промышленных комплексах, является актуальной задачей. Эта задача может быть решена только при активном участии морфологов, которым надлежит определить пределы адаптации к тому или иному фактору новых искусственно созданных условий обитания в промышленных комплексах, чтобы профилактировать структурные
изменения в организме животных, являющихся первым сигналом начала патологии органа (И.В.Хрусталёва, 1987, 1990).
Строение, рост и развитие трубчатых костей и их суставных, и метафизар-
ных хрящей крупного рогатого скота изучали Б.В.Криштофорова (1987), Л.Г.Харченко (1987), Х.Б.Баймишев, (1987), Б.П.Шевченко, Н.А.Малков, М.В.Андреев (1992). Объектом исследования служили кости грудных и тазовых конечностей плодов в возрасте 3, 6 и телят после рождения 3, 6 и 12 месяцев клеточного и пастбищного содержания (табл. 45).
Исследовали толщину компактного костного вещества, диаметр гаверсовых, фолькмановых каналов, остеонов, толщину перекладин и количество остеонов в одном поле микроскопа. Установлено, что толщина компактного костного вещества от плечевой, бедренной кости дистально вниз к пястным, плюсневым костям в плодном периоде развития уменьшается, а у телят после рождения, наоборот, от пястных, плюсневых костей вверх - возрастает.
45. Возрастные изменения компактного вещества костей конечностей
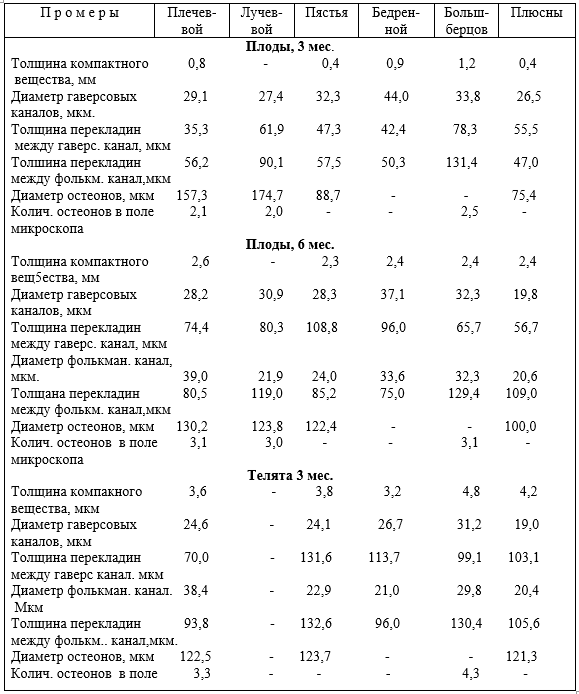
Диаметр гаверсовых и фолькмановых каналов значительно больше в плечевой кости, нежели – в пястных костях. Перекладины между гаверсовыми каналами в компактном веществе плечевой кости с возрастом утончаются, а в пястных костях – утолщаются. Толщина перекладин в каждом звене грудной конечности постепенно увеличивается. Это свидетельствует, что по звеньям, начиная с проксимального конца к дистальному концу грудной конечности, плотность компактного костного вещества возрастает
Рыхлость компактного вещества плечевой кости, относительно пястных костей, значительная и объясняется тем, что часть динамической нагрузки плечевой кости берут на себя мышцы плечевого и локтевого суставов. Тонус этих мышц уменьшает нагрузку на кость, что приводит к рыхлому строению костей.
При сравнении структурных показателей костей тазовой конечности выявлена следующая закономерность, что диаметр фолькмановых и гшаверсо-
вых каналов убывает в дистальном направлении конечности, но толщина перекладин увеличивается у плодов и телят в возрасте три месяца. Причем компактное костное вещество тазовой конечности толще, чем грудной от 2,2 до 3,6 раза. Установлено, что диаметр каналов с возрастом уменьшается, а толщина перекладин между ними увеличивается.
Данный анализ позволяет сделать вывод о неодинаковой функциональной значимости конечностей. Известно, что грудная конечность при стоянии и движении поддерживает переднюю часть тела над поверхностью почвы, а тазовая дополнительно выполняет локомоторную функцию, двигая туловище вперёд в горизонтальной плоскости. Таким образом, выполняет большую нагрузку, чем грудная конечность. Увеличение функциональной нагрузки на
кости тазовой конечности приводит к уплотнению и истончению компактного костного вещества. Приведённое выше предположение подтверждает морфологическое исследование бедренной кости северного оленя.
Известно, что крупный рогатый скот – «изнеженное животное», которое в зимнее время находится в помещениях, а северный олень ведёт полудикий образ жизни, круглый год, в полном смысле слова, находится под открытым небом и добывает себе корм сам «из под ноги». Сравнительный анализ структур бедренных костей этих животных показывает, что толщина компактного костного вещества бедренной кости северного оленя уступает толщине компактного вещества бедренной кости плодам и телятам крупного рогатого скота. Диаметр всех каналов костей северного оленя меньше, перекладины тоньше, чем крупного рогатого скота, но количество остеонов в поле микроскопа значительно больше (табл.46).
46. Структуры компактного вещества бедренной кости взрослых северного оленя и крупного рогатого скота
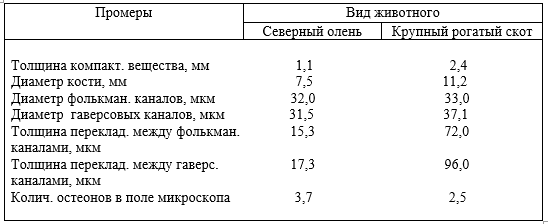
Из анализа следует, что компактное вещество бедренной кости оленя плотнее, а крупного рогатого скота – рыхлее. Полагаем, что на формирование костей северного оленя оказывают влияние суровые экологические условия обитания, добывания корма из под «ноги», генетически закреплённые в экономном расходовании органических и минеральных веществ при формировании костей как в утробе матери, так и после рождения. Рост трубчатых костей конечностей более активно развивается во втором полугодии после рождения телят, чем в первом. При сравнении роста костей в длину, выявлено, что при усиленной двигательной активности трубчатые кости тазовых конечностей опережает в росте длину грудных конечностей (табл. 47).
Из таблицы следует, что максимальный линейный прирост длины трубчатых костей приходиться на возраст 12 месяцев. Длина плечевой кости в этот промежуток времени увеличивается в 1,3, лучевой – 1,13, бедренной и большеберцовой, соответственно – в 1,17 и 1,17 раза. Массу наиболее интенсивно увеличивает лучевая кость – в 1,52 раза, далее бедренная и большеберцовая – 1,48 и плечевая – в 1,38 раза.
47. Длина трубчатых костей конечностей телят
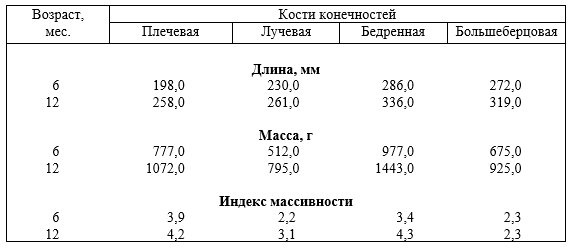
В абсолютных величинах эти показатели расположились следующем порядке: максимально прирост увеличивает плечевая кость – 50 мм, далее бедренная – 50 мм, большеберцовая – 47 мм и меньше всех лучевая – 31 мм. По приросту массы они распределились в следующей последовательности: лучевая – 283 г, плечевая – 295 г, большеберцовая – 300 г и бедренная кость 471 г. Отдельные исследователи для характеристики развития трубчатых костей используют индекс массивности (отношение длины к массе кости). Наименьший индекс массивности имеет большеберцовая кость. Несколько выше лучевая и бедренная, и наибольший – плечевая кость. В тоже время, изменение индекса массивности в костях происходит независимо от функции и положения костей в скелете конечностей, поэтому, мы считаем, что он полно не отражает рост и развитие костей. Различная степень двигательной активности больше влияет на рост, развитие костей конечностей телят. При этом генезис костей зависит от топографии кости в скелете конечностей.
Компактное вещество всех трубчатых костей конечностей у плодов в возрасте до трёх месяцев рыхлое, у плодов в возрасте шести месяцев становится уплотнённым, а у телят в три месяца после рождения - плотным и приобретает все черты, свойственные взрослым животным. Остеоны в костях конечностей могут располагаться в различных плоскостях, но чаще вдоль продольной оси кости. Это свидетельствует, что они лежат чаще в направлении силы действия тяжести и функции мышц на кость. Темп роста костей, в свою очередь, зависит от степени двигательной активности животного.
Гиперфункция вызывает рост массы, умеренный объём эпифизов, увеличение плотности остеонов в компактном веществе, усиливает гемодинамические процессы и рессорную функцию костей.
16. Толщина суставных хрящей в зависимости от двигательной активности
Исследование хрящей трубчатых костей молодняка крупного рогатого скота, находящегося в условиях различной двигательной активности, показало, что структура суставных хрящей изменяется с возрастом, и попадает в зависимость от положения кости в скелете конечностей. Наибольшая толщина суставных хрящей трубчатых костей отмечается у новорожденных телят. Исключение составляют хрящ большеберцовой кости, наибольшая толщина которого на проксимальном эпифизе отмечается в возрасте 18 месяцев, а дистального – в шесть и проксимального эпифиза лучевой кости - в 18 месяцев (табл. 48).
48. Толщина суставных хрящей трубчатых костей конечностей с возрастом
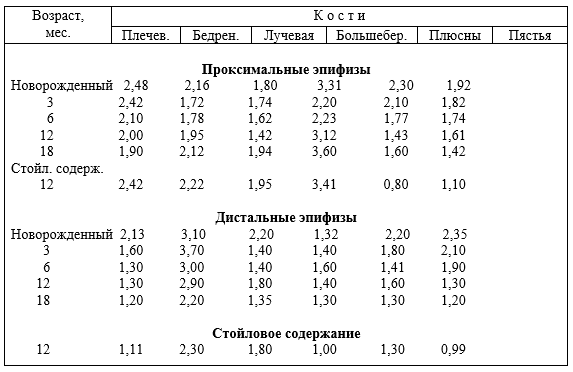
У новорожденных телят толщина хрящей проксимальных эпифизов костей грудной конечностей последовательно уменьшается от плечевой кости к лучевой и пястным костям. Тазовой конечности, соответственно от бедренной кости - к плюсневым костям. С возрастом суставной хрящ равномерно истончается на плечевой и пястных костях, на бедренной и большеберцовой возрасту телят три месяца несколько снижается, а к 18 месяцам вновь повышается, соответственно - в 1,23 и 1,64 раза.
Суставной хрящ плечевой кости истончается до 12 месяцев, а в 18 – вновь увеличивается. Толщина хрящей дистальных эпифизов новорожденных телят убывает в последовательности: от бедренной, к пястным, плюсневым, лучевой, плечевой и большеберцовой костям. К 18-месячному возрасту происходит равномерное истончение суставного хряща плечевой, бедренной и пястных костей. Из этой обоймы выпадает лучевая кость, её суставный хрящ уменьшается в толщине у телят в шесть месяцев, в 12 - незначительно возрастает, а к 18 месяцам вновь - уменьшается. Что касается суставного хряща дистального эпифиза большеберцовой кости, его толщина к возрасту телят шесть месяцев возрастает в 1,21 раза, а к 18 месяцам – истончается до толщины хряща новорожденных.
Сравнение толщины суставных хрящей трубчатых костей молодняка крупного рогатого скота, с различной функциональной нагрузкой на скелет конечностей, показывает, что хрящ проксимальных эпифизов трубчатых костей конечностей телят клеточного содержания был толще, а телят пастбищного содержания - тоньше. Так, плечевой – в 1,44; бедренной – 1,14; лучевой – 1,37; большеберцовой – 1,09 раза, а плюсневых - в 1,79 и пястных – 1,46 раза. Что касается дистальных концов исследуемых костей, то толщина суставных хрящей телят пастбищного содержания превышала таковой у телят стойлового содержания: плечевой – 1,17; бедренной – 1,04; большеберцовой – 1,40; плюсневых – 1,23; пястных – в 3,1 раза.
И так, различная степень двигательной активности телят влияет на рост, развитие суставных хрящей. При этом развитие хрящей зависит от топографии кости в скелете конечности. При гиперфункции на конечности суставные хрящи
проксимальных эпифизов истончаются, а дистальных - утолщаются. Различная степень толщины хрящей дистальных эпифизов относительно проксимальных зависит от суставного угла и передачи силовых толчков, приходящихся на кости.
Вопросы для контроля
17. Кровоснабжение костей
Относительная масса скелета человека составляет 8 – 10%, свиней 6 – 9%, коз 7 – 9%, овец в зависимости от породы 9 – 15%, новорожденных симментальского скота – 24,4%, 1,5-годовалого молодняка – 11,3%, взрослого скота – 10,4%, лошадей – 13% от массы тела (В.Н.Жеденов, 1965).
Необходимо помнить, что кость – это живая ткань, которая при динамических нагрузках накапливает в себе продукты метаболизма, в связи с этим, появляется усталость костной ткани. Кость подвержена различным заболеваниям – остеомаляции, остеодистрофии, остеопорозу и др. Б.Фолков, Э.Нил (1976) указывают, что обменная активность и кровоснабжение костной ткани значительны, особенно, в период роста и во время заживления переломов костей.
В полостях костей заключён паренхиматозный красный и жёлтый мозг, количество которого у собак может достигать 950 г, овец - 1,2 кг, свиней 1,7 –
2,5 кг, крупного рогатого скота 6,03 - 7,75 кг, лошади - 9,5 кг, а у жеребцов и более. У домашних животных объём костного мозга изучен недостаточно, а функциональное его значение велико. Гемопоэз идёт в красном костном мозге, но при больших потерях крови, анемии функцию кроветворения может выполнять и жёлтый мозг.
Сосуды кости, костного мозга тесно связаны как между собой, так и с сосудами окружающих тканей, органов посредством анастомозов и коллатералей. По косвенным данным кровоток в кости крупного рогатого скота достигает 48–72 мл в минуту на 100 г костной ткани. Если масса свежей плечевой кости взрослого скота достигает в среднем 1200 г, не трудно подсчитать, что через неё проходит 576 – 864 мл крови в минуту, или в среднем – около 720 мл. Это много. Не совсем точные данные показывают, что кровоток кости высокий и обеспечивает костную ткань, и мозг не только питательными, и минеральными веществами, но и выносит из них созревшие форменные элементы крови и отработанные продукты.
Стенка артерий костного мозга тонкая. В стенке артериол разреженно встречаются гладкие мышечные клетки. При отхождении от артериол прекапилляров и от последних – капилляров, в соустье их имеется сфинктеры, состоящие из нескольких гладких мышечных клеток. Капилляры чаще в красном мозге и реже – в жёлтом, расширяясь, переходят в синусоиды (рис.11).

Рис.11. Со-устьевый сфинктер. Импрегнация
азотнокислым серебром. Об.90, ок. 7.
Синусоиды – это истинные «обменные звенья» костного мозга, где кровоток резко замедлен в связи с расширением до 60 мкм. Если площадь сечения истинного капилляра крупного рогатого скота достигает в среднем 20 – 26 мкм2, то синуса 188 – 210 мкм2. Стенка его тонкая, просвет широкий, боковое давление и скорость движения крови низкие. Стенка синусоида контактирует с жидкостным пространством и с ячеями ретикулярной основы костного мозга. По
этому, жидкая среда мозга и компактное вещество трубчатых костей, сдерживают переполнение сосудов кровью и предохраняют их от разрыва.
Стенка синусоида представлена эндотелиальной выстилкой и тонкой прослойкой рыхлой соединительной ткани (экстерны). На границе между эндотелиальными клетками имеются межклеточные щели, через которые, из ячей заполненных регидной жидкостью, в кровеносное русло заносятся созревшие форменные элементы крови.
Симпатическая часть вегетативной нервной системы влияет на кровообращение в сосудах кости как при переломах, так и при их удлинении (Л.Л.Абрамова, Н.И.Антонов, 1999; Н.И.Антонов, Л.Л.Абрамова, 1999). Однако путём «почкования» могут формироваться как новые сосуды, так анастомозы и коллатерали. По этому, действие симпатической части нервной системы может существенно влиять на сосуды кости и мозга, и на их кровоснабжение (рис.12).
Кровоток в костной ткани и мозге всегда повышается при гипоксии, анемии и потери крови.

Рис.12. Иннервация кровеносных сосудов.
Импрегнация азотнокислым серебром. Об. 20, ок.7.
18. Особенности кровоснабжения трубчатой кости
Что бы разобраться в особенностях кровоснабжения трубчатых костей, надо понять строение и деление кости на составные части. Любая трубчатая кость состоит из тела – диафиза. Тело вверх и вниз переходит в метафизы, которые посредством хрящей, фиксируются с апофизами (с костными выступами костей. буграми, отростками) и эпифизами – суставными концами трубчатых костей.
Тело кости представлено компактным костным веществом, а эпифизов и метафизов – губчатым веществом, в ячеях которого находится красный мозг.
Снаружи кость покрыта надкостницей, а костная полость выстлана эндоостом и заполнена жёлтым костным мозгом.
Компактное костное вещество состоит из генеральных пластинок, остеонов. Остеоны, в свою очередь, состоят из циркулярных пластинок, в центре которых располагается гаверсов канал. Остеоны соединены между собой вставочными пластинками. Со стороны надкостницы в компактное костное вещество проходят фолькмановы каналы, соединяющиеся с гаверсовыми. Чаще на каудальной (пальмарной и плантарной) поверхности диафиза костей располагается два – четыре внутренностных (питательных) отверстия, через которые в полость кости
проходят крупные артерии, нервы и выходят вены. Вокруг суставных поверхностей эпифизов располагается большое число (54 - 68) отверстий, диаметром 0,4 – 1,8 мм. Через крупные диафизарные отверстия в костное губчатое вещество проходит артерия, нерв и выходит вена. Через отверстия, диаметром 0,4 – 1,0 мм, выходят вены, но в гиалиновом хряще сосудов нет. Они располагаются в базальной зоне между хрящом и губчатым веществом, и обеспечивают питание хряща, кости и кальцинирование матрикса (В.Н.Павлова, 1980). Под гиалиновым хрящом эпифизарные артерии, анастомозируя, образуют артериальные терминалии.
Диафизарные сети образуются артериями надкостницы, проходящие через
фолькмановы каналы, а со стороны костной полости ветви главной внутренностной артерии идут в губчатое вещество метафизов, соединяясь между собой, образуют метафизарную сеть красного мозга. Из этой сети вены, диаметром 0,4 – 1,0 мм, выходят через мелкие эпифизарные отверстия.
Рядом, проходящие с трубчатой костью магистральные артерии, направляют ветви как в окружающие кость органы (мышцы, сухожилия, фасцию), так и в надкостницу и кость. В надкостнице они образуют микроциркуляторную сеть, из которой в компактное костное вещество через фолькмановые каналы вместе с артериями проходят нервы. В итоге, в компактном веществе формируется сосудистая и нервная сети, обеспечивающие питание кости и иннервацию. Из этого следует, что кость – это живая ткань, которая имеет сложную собственную сосудистую, нервную сети. В этих сосудистых сетях кости при гиперфункции
накапливаются продукты метаболизма, вызывающие её усталость, а нервные сети передают этот сигнал в подкорковые и корковые центры. Поэтому костная ткань как и другие органы нуждается в отдыхе (рис. 13).
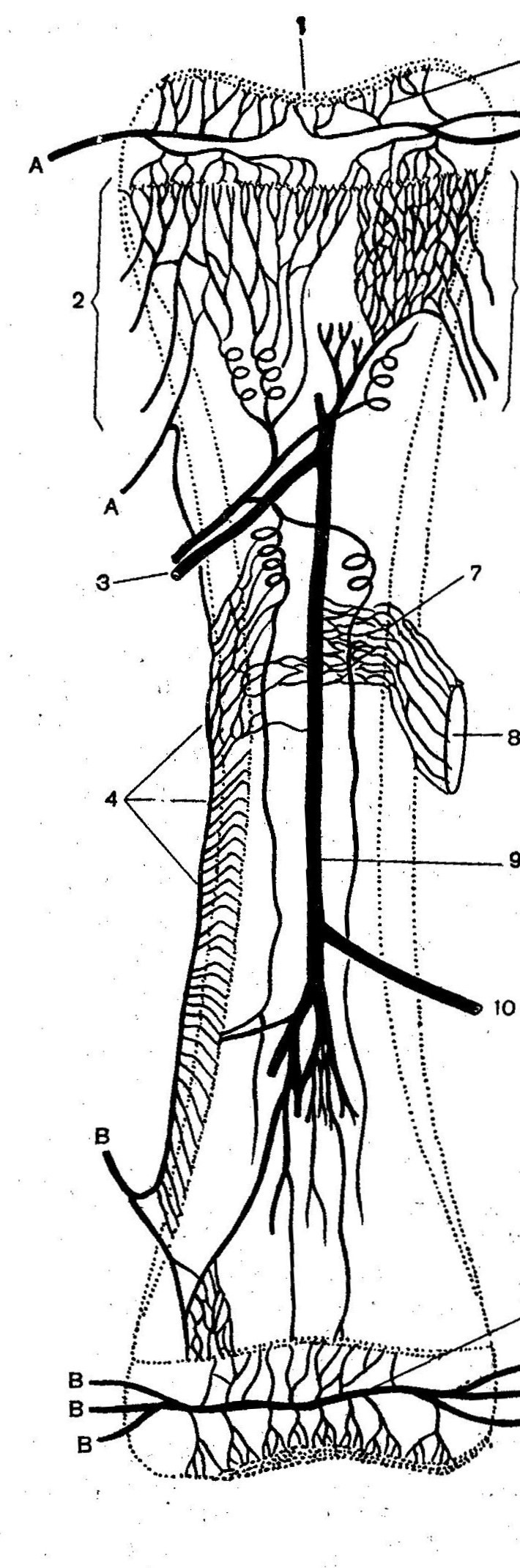
Рис. 13. Схема кровоснабжения трубчатой кости. 1 – суставной хрящ; 2 – метафизарная сеть; 3 – питающие артерия и вена; 4 –микроциркуляторное русло надкостницы; 5 – субхондральная сосудистая сеть эпифиза; 6 – синусоиды и метафизарные вены; 7 – синусоиды костного мозга диафиза; 8 – межорганные сосуды; 9 – диафизарная вена; 10 – большая вена эмиссария; А – артерии, В – вены (по Б.Фолкову, Э.Нил, 1976).
Главная питающая (внутреностная) артерия в красном и жёлтом костном мозге образует сложную сеть, состоящую из капилляров и синусоидов. Из этой сети кровь выносят вены выпускницы (эмиссарии), которые могут находиться в триаде (артерия, нерв, вена) питательного отверстия, или выходить отдельно.
И так, в длинных трубчатых костях артерии формируют: периоссальные сети,
эпифизов и апофизов, диафизов и метафизов, наконец, красного и жёлтого костного мозга. Из этого следует, что кровеносные сети трубчатых костей тесно
связаны между собой посредством анастомозов и коллатералей, взаимообусловлены строением кости, её положением в звене скелета конечностей и изменяются при динамических нагрузках на кость.
Вопросы для контроля
- Где располагается красный и жёлтый костный мозг?
- Особенности строения костных кровеносных сосудов?
- Что такое синусоиды костного мозга?
- На какие суставные части делится трубчатая кость?
- Строение трубчатой кости – как органа?
- Объяснить процесс кровоснабжения костной ткани?
- Какие сети образуют питательные артерии в кости?
19. Особенности вен трубчатой кости
Главная вена эмиссария костей стопы по месту выхода и величине диаметра непостоянная (Б.В.Криштофорова, 1988). В случае присутствия обнаруживается на каудальной поверхности нижней трети трубчатых костей. На плечевой кости она имеет диаметр 0,6–1,2 мм, в 42% случаев может отсутствовать, на бедренной кости она более постоянна (рис.13).
Внутренностная вена костей предплечья по месту выхода из кости более постоянная, имеет больший диаметр, чем плечевой кости, который позволяет ей выводить кровь из тела и мозга трубчатых костей. В остальных костях она слабо развита, имеет диаметр 0,3 – 0,6 мм, и не отражает истинный размер диаметра вены отводящей кровь из кости. Толщина стенок вен при выходе из отверстия кости достигает 140 мкм, в соустье с другими венами - 240 мкм.
Таким образом, большая часть вен плечевой, бедренной кости выходит из отверстий эмиссарий, расположенных по краю суставных поверхностей эпифизов, бурс, синовиальных влагалищ, т.е. там, где они больше защищены от компрессионных действий сухожилий, мышц, связок.
Артерии и вены тонкостенные, но артериолы снабжены миоцитами и компрессионными нервными волокнами. Б.Фолков, Э.Нил (1976) указывают, что перед участком, где исчезают гладкие мышечные клетки, есть сфинктероподоб-
ные устройства, которые обозначают границу перехода артериолы в прекапилляр или ответвления прекапилляров и капилляров от артериол.
Капилляры, расширяясь, могут переходить в синусоиды костного мозга. Они редко, но встречаются и в жёлтом мозге. Там, где имеются синусоиды, располагаются вокруг них обходные кровеносные пути. Стенки синусоидов тонкие (см.гл.17), их просветы контактируют через межклеточные эндотелиальные щели с жидкостью ретикулярной основы костного мозга, где концентрируются созревшие кровяные клетки. В итоге, создаются условия обмена между внутрисинусоидным и межсинусоидным пространствами, через которые обеспечивается перемещение созревших клеток.
Вены тонкостенные и имеют большой диаметр. Они не снабжены контрактильными элементами и не имеют иннервации (Б.В.Криштофорова, 1988). Стенка их по строению похожа на стенку капилляра. Рыхлой наружной оболочкой вены прикрепляются к ретикулярной основе костного мозга, с артериями и костными перекладинами. Объём сосудистого ложа кости достигает
10% объёма ткани. В просвете вен костного мозга, эпифизов, метафизов и диафизов встречаются структуры подобные клапанам, особенно, в местах изменения направления магистрального ствола. В этих местах встречаются складки, напоминающие одностворчатые клапаны, трабекулы, диаметром 45–100 мкм, внутривенные подобные связкам образования (трабекулы). Артерии, диаметром до 270 мкм, могут проходить внутри просвета вен или через вены, как бы прободая их. После выхода из вены могут формировать паравенозное сплетение. Все эти образования играют большую роль в гемодинимике кости.
Притоковые венозные ветви проксимального метафиза, сливаясь, формируют диафизарную костномозговую вену. Опускаясь дистально, она отдаёт, как правило, вену выпускницу на границе средней и нижней трети трубчатой кости, а её основной ствол идёт вниз по внутренней поверхности полости кости и принимает дополнительно притоковые вены костного мозга, и выходит из полости через многочисленные выпускные отверстия дистального эпифиза трубчатой кости.
Таким образом, венозное русло красного и жёлтого костного мозга изучено
ещё недостаточно. Имеющиеся литературные данные указывают на сложность строения и формирование его русла, которое влияет на гемопоэз и гемодинамику кости. При этом, венозное русло трубчатых костей как наиболее пластичное, воспринимая механоресорные воздействия костей, тоже начинает перестраивать-ся.
Вопросы для контроля
- Какие особенности строения внутрикостных вен?
- Какие встречаются структуры в стенке костных вен?
- Укажите внутривенные образования, в просвет костных вен?
20. Артериальная магистраль грудной конечности
Представлена подключичной, подмышечной, плечевой, срединной артериям
В области кисти её формируют глубокая пястная пальмарная артерия, являюща-
яся продолжением дистально лучевой артерии, глубокая пястная пальмарная латеральная артерия - продолжение на кисть каудальной межкостной артерии и поверхностная пястная пальмарная артерия, которая является непосредственным продолжением на кисть срединной артерии. Они в области пястья образуют проксимальную и дистальную глубокие дуги (рис. 14).
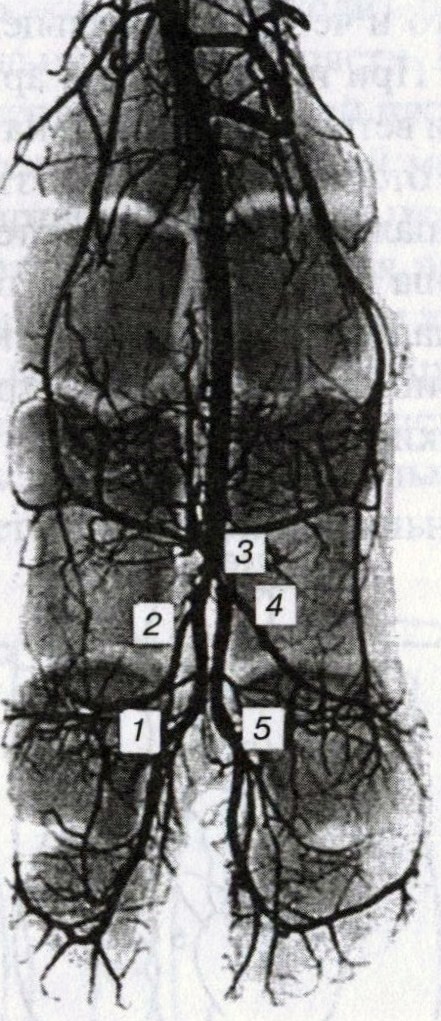
Рис. 14. Рентгеновый снимок артерий фаланг пальцев
грудной конечности. 1, 5 – специальные осевые артерии
III-IV пальцев; 2, 4 – артерия мякиша III-IV пальца:
3 – межпальцевая пальмарная артерия (по Н.С.Иванову, 2009).
Непосредственно на пальцы из глубокой дистальной пальмарной дуги выходит межпальцевая пальмарная артерия, разветвляющаяся на специальные осевые артерии III–IV пальца. По ходу они отдают артерии первой, второй фаланги, к мякишам и переходят в венечные артерии, образующие в венечном канале третьей фаланги терминальную (концевую) дугу. Концевой дуги через многочисленные эмиссарные отверстия выходят артериальные ветви, обеспечивающие кровоснабжение париетальной и подошвенной поверхностей копытца крупного рогатого скота.
21. Вены грудной конечности
Вены грудной конечности формируют глубокую и поверхностную магистрали. Глубокая венозная магистраль конечности сопровождает ход и ветвление артерий, и нервов, и через подключичную вену выносит кровь в краниальную полую вену, а последняя открывается в правое предсердие сердца. Она выходит из глубокой пальмарной дуги, которую образуют общие дорсальные и пальмарные I–IV пальца вены.
Поверхностная венозная магистраль выходит из дорсальной общей пальцевой вены, ветвь которой переходит проксимально в подкожную головную вену. Перед тем как перейти в подкожную головную вену они участвуют в бразовании с общими пальмарными (плантарными) межпальцевыми венами глубокой дуги,
из которой выходит глубокая венозная магистраль. Из этого следует, что из фаланг пальцев кровь может оттекать как в поверхностную венозную магистраль, так и в глубокую. Из краткого анализа следует, что внутрикостные артерии и вены фаланг пальцев исследованы крайне слабо.
Вопросы для контроля
- Перечислите артерии, составляющие магистраль грудной конечности?
- Какие артерии кровоснабжают пальцы и копытца?
- Какими венами представлена глубокая и поверхностная венозная магистраль грудной конечности?
- Откуда выходит и куда открывается глубокая венозная магистраль грудной конечности?
- Откуда выходит и куда открывается поверхностная (подкожная) венозная магистраль грудной конечности?
22. Артериальная магистраль тазовой конечности
Включает наружную подвздошную, бедренную, подколенную, переднюю и заднюю большеберцовые артерии.
Передняя большеберцовая артерия крупного рогатого скота выходит на спинковую поверхность заплюсны и плюсны - как дорсальная артерия стопы. В области заплюсны получает прободающую заплюсневую артерию и отдаёт проксимальную и дистальную прободающие плюсневые артерии на подошвенную поверхность стопы. Далее она переходит в начале в общую дорсальную III-IV пальца артерию, которая отдаёт дорсальные специальные пальцевые артерии, а сама проходит через межпальцевое пространство на подошвенную поверхность стопы и вступает в анастомоз с глубокой плантарной дугой.
Задняя большеберцовая артерия выходит на каудальную поверхность голени и кровоснабжает в основном мышцы разгибатели заплюсны, плюсны и фаланг пальцев, концевой её ветвью является медиальная лодыжковая артерия.
Артерия сафена отходит от бедренной артерии на уровне средней трети бедра. Проходит дистально в начале по медиальной поверхности бедра, затем выходит заднюю поверхность голени с одноименным нервом, веной. Опускаясь дистально, идёт по краниомедиальной поверхности сухожилия поверхностного сгибателя пальцев и трёхглавой мышцы голени (ахиллово сухожилие). Ниже пяточной кости она делится на латеральную и медиальную плантарные плюсне-
вые артерии, которые идут вниз – как специальные плантарные артерии III-IV пальца. Последние переходят в венечные артерии, которые в области III фаланги образуют, по подобию копытца грудной конечности, терминальную дугу (см. рис.14). Из терминальной дуги через отверстия эмиссарии выходят артериальные ветви в кожу подошвенной поверхности копытца.
23. Вены тазовой конечности
Вены тазовой конечности, как и грудной, образуют две магистрали – глубокую и поверхностную.
Глубокая венозная магистраль соответствует ходу, ветвлению артерий, нер-
вов тазовой конечности. Поверхностная (подкожная) венозная магистраль представлена латеральной и медиальной подкожными венами (сафена).
Латеральная вена сафена образуется слиянием краниальной и каудальной ветви. Краниальная ветвь выходит из поверхностной дорсальной дуги, образованной слиянием общих дорсальных вен III-IV пальца. Последние сформированы слиянием собственно дорсальных вен III-IV пальца. Поверхностная дорсальная дуга соединяется с плантарной дугой. Каудальная ветвь вены сафены выходит из анастомоза задней ветви медиальной вены сафены. Из этого следует, что кровь из копытца может оттекать вверх - к сердцу как через латеральную вену сафену, так и через медиальную.
Краниальная и каудальная ветви, объединившись в один ствол, образуют латеральную подкожную вену сафену, которая, как правило, открывается в окружную медиальную вену бедра. Редко может самостоятельно открываться в бедренную вену выше коленного сустава.
Медиальная вена сафена формируется слиянием латеральной и медиальной плантарных плюсневых вен.
Латеральная плантарная вена выходит из проксимальной глубокой плантарной дуги, последняя сформирована плантарными плюсневыми венами III–IV пальца. Через них она соединяется с дистальной глубокой дугой. В образовании последней принимают участие общие плантарные вены III–IV пальца и дистальная прободающая ветвь. Медиальная планатрная вена образована тоже слиянием двух ветвей. Менее развитая глубокая ветвь, выходит из проксимальной глубокой плантарной дуги, более развитая поверхностная ветвь является продолжением общих плантарных плюсневых вен III–IV пальца. Последние являются продолжением специальных плантарных вен пальцев. Чаще изменяющиеся при гиподинамии.
Из краткого анализа следует, что внутрикостные вены фаланг пальцев, как и кости пальцев грудной конечности крупного рогатого скота, изучены поверхностно. Одновременно – это звено чаще подвергается структурному изменению при гиподинамии.
Вопросы для контроля
- Составьте артериальную магистраль тазовой конечности?
- Откуда выходит и где располагается артерия сафена?
- Какими венами представлена глубокая венозная магистраль тазовой конечности?
- Чем представлена поверхностная (подкожная) венозная магистраль)
- Откуда выходит латеральная вена сафена?
- Откуда выходит медиальная вена сафена?
24. Скелет и вены пальцев конечностей
Скелет пальцев грудной и тазовой конечностей представлен первой (проксимальной), второй (средней) и дистальной (копытцевой) фалангами. Первую и вторую фалангу следует отнести к коротким трубчатым костям, а третью – к губчатым. Проксимальная, средняя фаланги по строению похожи друг на друга, только вторая короче первой. Грудной конечности они толще и короче, чем тазовой. В поперечном сечении они имеют полуовальную форму, слегка сплющенную в латеромедиальном направлении (рис. 15).
На фалангах различают основание (суставную ямку), тело и головку (блок). Суставная ямка направлена вверх, а головка вниз и вперёд. Копытцевые кости крупного рогатого скота асимметричные, имеют в сагиттальной плоскости
треугольную форму. Венечным (подошвенным) краем копытце делится на париетальную и подошвенную поверхности. В крае располагается венечный (подошвенный) канал, в котором находится терминальная кровеносная дуга. Канал большим числом эмиссарий (отверстий) открывается на париетальной и подошвенной поверхностях. Через них входят артериальные ветви в кожу копытца и выходят венозные ветви в терминальную дугу.
Рис. 15. Акроподий задней конечности.
1 – плюсневые кости; 2 – проксимальный, 3 - средний,
и 4 – дистальный фаланг (копытцевая кость). Бык месячного возраста.
Основными зонами выхода вен эмиссарий из копытцевой кости являются париетальная и подошвенная поверхности. Париетельную поверхность делят на аксиальную – осевую (межкопытцевая) и абаксиальную – неосевую или наружную поверхность. По краю суставной поверхности насчитывается более 36 отверстий эмиссарий, на париетальной – до 58, на подошвенной – более 28 отверстий, по подошвенному краю – 76. Медиально от суставной поверхности находится аксиальное входное отверстие венечного канала, а латерально, через разгибательный отросток, лежит абаксиальное проксимальное и абаксиальное дистальное отверстия (рис. 16).

Рис.16. Париетальная поверхность копытцевой кости
1У пальца. 1 – суставная поверхность; 2 – неосовое отверстие;
4 – отверстия эмиссарии венечного канала. Бык 9 месяцев.
На подошвенной поверхности копытца различают 2-4 аксиальных отверстия, ведущих в подошвенный канал и четыре – пять абаксиальных отверстий. Наличие такого количества входных и выходных отверстий свидетельствует об интенсивности кровоснабжения копытца, которое, в свою очередь, зависит от двигательной активности животного (рис. 17). При этом выходные отверстия их имеют «лыски», предохраняющие сосуды от травм.
Рис. 17. Плантарная поверхность копытцевой кости
1У пальца. 1 - отверстия эмиссарии подошвенного канала
располагаются в сосудистом жёлобе.
Терминальная дуга, после выхода из осевого и неосевого отверстий подошвенного канала, соответственно, переходит в III–IV осевые и неосевые собственные (специальные) пальмарные - ладонные (плантарные - подошвенные) пальцевые вены, выносящие кровь из губчатого вещества копытец. Последние, открываются вены, выносящие кровь из венчика копытца, средней и проксимальной фаланги (рис.18).
Рис.18. Венозный коллектор венчика копытца.
1 – венозная сеть основы кожи париетальной поверхности
копытца; 2 – венозный коллектор венчика копытца;% 3 –
париетальная венозная сеть копытца. Бык 14 месяцев.
Из их слияния выходит III-IV общие пальмарные (плантарные) пальцевые вены, которые в начале образуют поверхностную дорсальную дугу, из последней выходит латеральная вена сафена (см.гл.23), а затем глубокую дистальную пальмарную (плантарную) дугу, из этой дуги выходят III–IV пальмарные (плантарные) пястные (плюсневые) вены, вливающиеся в проксимальную глубокую дугу. Из этой дуги грудной конечности выходит глубокая венозная магистраль, а тазовой конечности - медиальная вена сафена и глубокая венозная магистраль (рис.19, 20).
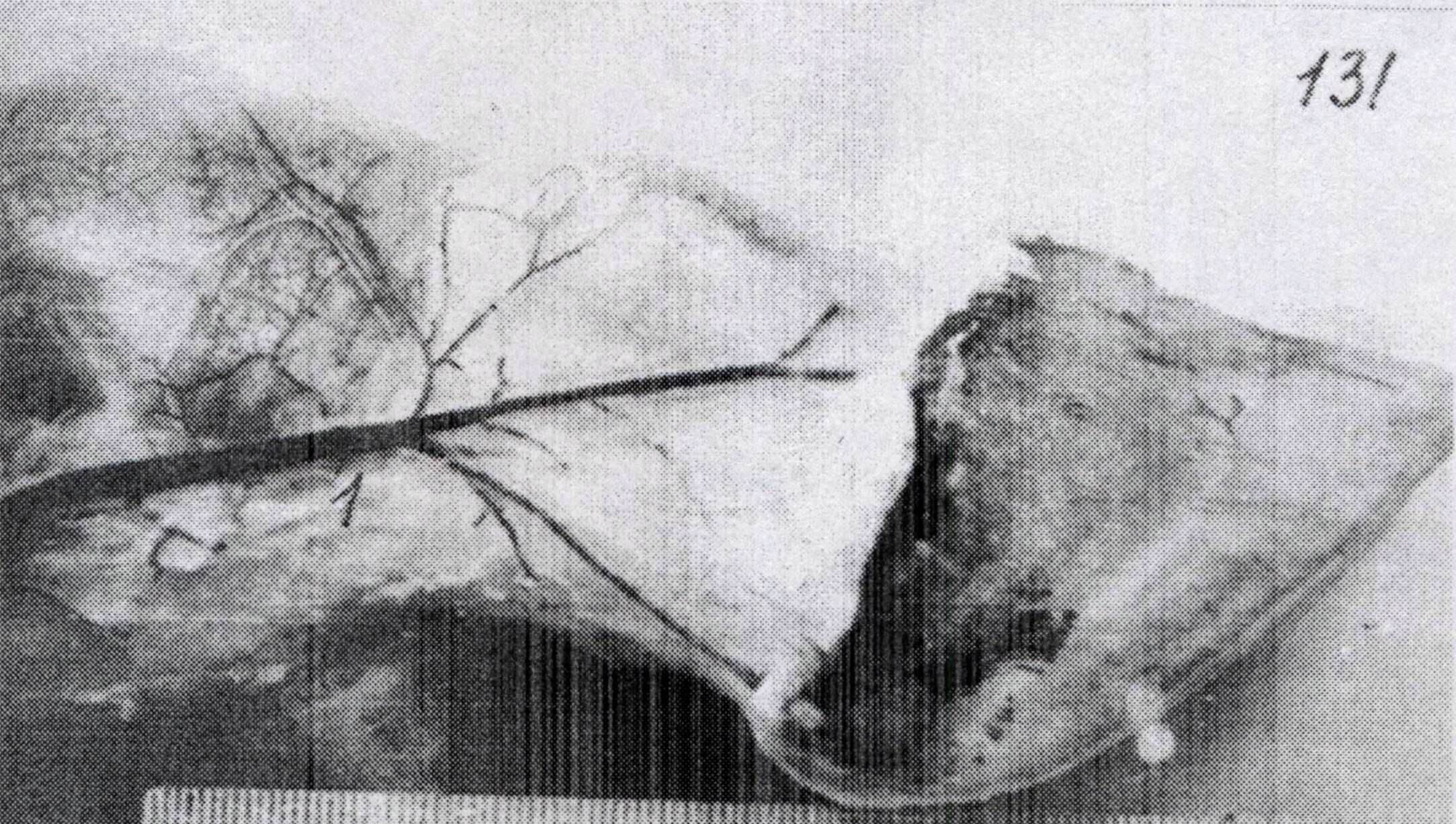
Рис. 19. Обшая плантарная медиальная вена Ш пальца.
1 – осевая собственная планатьарная вена Ш пальца.
Бык четыре месяца.
Необходимо отметить, что у лошади хорошо изучены как внутрикостные, так внекостные вены копытной кости (А.Ф.Климов, 1955). Но копытца крупного и мелкого рогатого скота исследованы слабо указывают В.Б.Борисевич (1982), Х.Б.Баймишев (1984), Н.С.Иванов (2006), а поражения копытцев жвачных встречается чаще, чем лошадей.
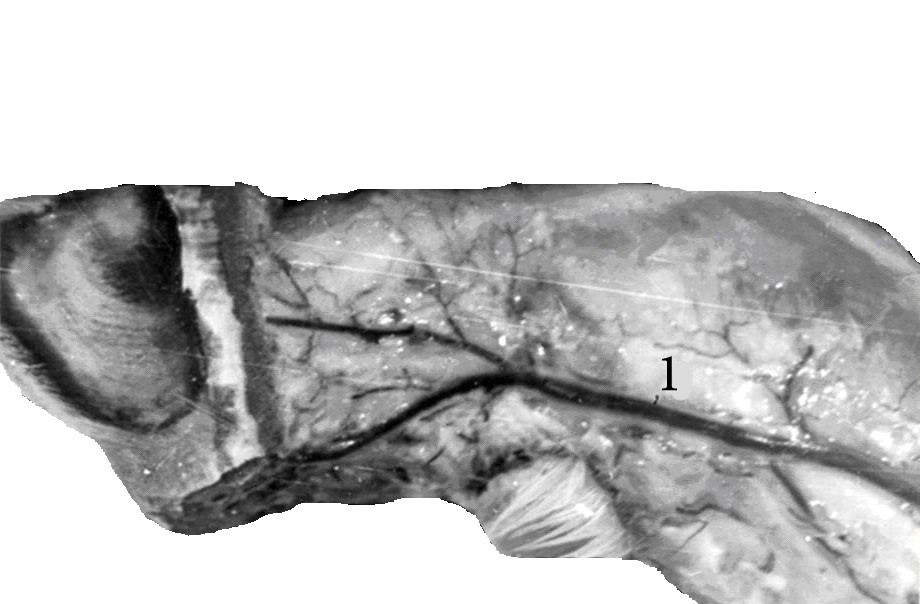
Рис. 20. Общая плантарная латеральная вена IV пальца.
1 – неосевая собственная плантарная вена IV пальца.
Бык четыре месяца.
В настоящее время большое значение придаётся изучению воздействия неблагоприятных факторов окружающей среды и условий промышленных комплексов на скелет пальцев и их сосуды. И.В.Хрусталёва, Э.И.Воробьёва, В.Н.Павлова (1987) пишут: «Одна наиболее из актуальных задач, которая стоит сейчас перед животноводами, выяснение оптимальных условий содержания животных в условиях промышленных комплексов. Эта задача может быть решена только при активном участии морфологов, которым надлежит определить пределы адаптации к тому или иному фактору новых искусственно созданных условий обитания в промышленных комплексах, чтобы профилактировать появление деструктивных изменений в организме продуктивных животных, являющееся первым сигналом начала патологии органа».
В.Б.Борисевич (1982), Х.Б.Баймишев (2002) отмечают, что чаще в промышленных комплексах травмируются копытца наружных пальцев, а в местностях с близким расположением вод к поверхности почвы, в скученных
сырых помещениях для скота, развивается копытная гниль (некробактериоз) и др. В связи с чем, резко понижается продуктивность животных.
Известно, что в области кисти и стопы располагается сложный связочно- сухожильный комплекс с синовиальными бурсами, влагалищами, сезамовидными костями, которые не только влияют на ход, ветвление сосудов, но и на кровоток в фалангах пальцев, особенно во время гиподинамии и гипердинамии. Поэтому, в качестве исходного исследовательского материала взяты были пальцы задней конечности, крупного скота, содержащегося в условиях промышленного комплекса. Изучены внутрикостные вены пальцев – как наиболее пластичное звено кровеносной системы, чаще подвергающиеся изменению структуры стенки и просвета вен при различных динамических нагрузках.
25. Вены париетальной стенки копытца
Копытце делится на кайму, венчик, париетальную и подошвенную поверхности. В области каймы и венчика кожа представлена эпидермисом, основой кожи и подкожным слоем, париетальной и подошвенной поверхностей – эпидермисом и основой кожи, подкожного слоя нет.
Стенка роговой капсулы копытца состоит из глазури, трубчатого и листочкового слоёв. Эпидермис каймы производит глазурь, венчика – трубчатый, а стенки – листочковый слои рога стенки копытца. Мякиш располагается в заворотном углу копытца, позади сустава третьей фаланги, сформирован – эпидермисом, основой кожи и подкожным слоем и выполняет осязательную
функцию. На париетальной, подошвенной поверхностях основа кожи соединяется с надкостницей III фаланги. Эпидермис основы кожи подошвеной поверхности производит роговой слой копытца (рис.21).
Рис.21. Копытца 14 – месячных бычков. Левое
бычка клеточного содержания, правое, получавшего ДПД.
Методами послойного тонкого препарирования под микроскопом МБС-2, на просветлённых и коррозионных препаратах установлено, что на латеральной стенке париетальной поверхности копытца венозная сеть представлена 3–4, а на медиальной 2 - 3 слоями. В области венчика, соответственно, три и четыре.
В листочковом слое латеральной стенки париетальной поверхности копытца находится тонкая венулярная сеть, в основе кожи – крупная петлистая венозная сеть, которая сливается в вены, диаметром 0,7–1,2 мм, направляющиеся от подошвенного края вверх в коллектор венчика копытца. Третья сеть надкост- ницы копытца представлена всеми звеньями микроциркуляторного русла, а области венчика дополнительно находится подкожная венозная сеть, из которой кровь оттекает в венозный коллектор копытца.
В листочковом слое и основе кожи медиальной стенки париетальной поверхности копытца сеть представлена венами, диаметром 0,12–1,1 мм, состоящая из крупных, разнообразной формы петель. Первые две сети соединяются тесно с сетью надкостницы третьей фаланги. Сосуды этой части копытца открываются тоже в венозный коллектор венчика. В эту венозную сеть через отверстия эмиссарии открывается более 76 внутрикостных вен, диаметром 0,12–1,6 мм, выносящих кровь из губчатого вещества третьей фаланги. Из кoллектора венчика копытца кровь оттекает в общую дорсальную вену III или
IV пальца и в медиальную вену III и в латеральную IV пальца (рис. 22).

Рис. 22. Венозная сеть основы кожи боковой стенки
копытца. Просветлённый препарат, бык четыре месяца.
Коллектор венчика III-го копытца бесклапанным анастомозом соединяется с коллектором венчика IV-го копытца, выполняя с одной стороны функцию коллектора и гидравлической подушки, а с другой – насоса, которая способствует оттоку крови из копытца вверх. Так как внутрикостные вены имеют тонкую стенку и слабо развитую среднюю оболочку, кровь вверх она протолкнуть не может. Механизм оттока заключается в следующем. При опоре конечности о поверхность почвы, кровь из коллектора венчика копытца под тяжестью тела выдавливается в дорсальные и пальмарные (плантарные) общие пальцевые вены. При поднятии конечности, в связи с тем, что внутри лежит жёсткая стенка копытцевой кости, наружу – такая же жёсткая роговая стенка венчика копытца, а между ними в основе кожи находится венозный коллектор, поэтому кровь из венозной париетальной и подошвенной кожной сети копытца устремляется в коллектор по принципу - где ниже давление. В обратном направлении при опоре о почву кровь течь не может, так как в соустьях вен с коллектором находятся двухстворчатые клапаны.
Диаметр отводящих вен суставного конца и подошвенного края копытца с возрастом животных увеличивается в зависимости от степени двигательной активности животных. Так, диаметр вен эмиссарий копытца новорожденного телёнка равен 0,5 мм, в месячном возрасте телят клеточного содержания 0,62 мм, четырёхмесячном – 1,03 мм, девятимесячном – 1,14 мм и в 14-месячном – 1,3
мм, париетальные сети имели разреженные ячеи (рис.23). Животные, получав-
шие дополнительное принудительное движение (ДПД), в месячном возрасте имели диаметр вен в 1,26 раза и в старших возрастах, соответственно – в 1,20; 1,23 и 1,31 раза больше (табл.48, рис. 24).
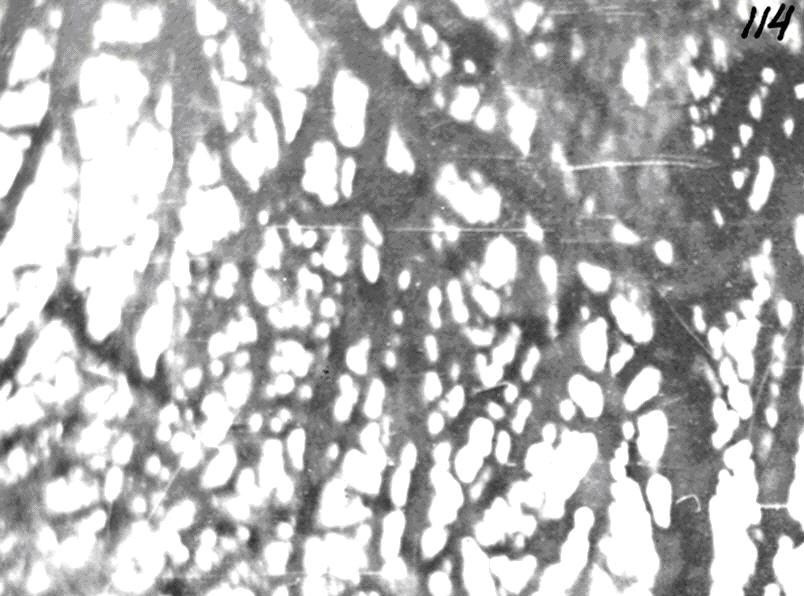
Рис. 23. Разреженные венозные сети основы кожи париетальной
поверхности копытца при гиподинамии. Контроль, бык 14 месяцев
48. Морфометрия вен копытцев тазовой конечности бычков
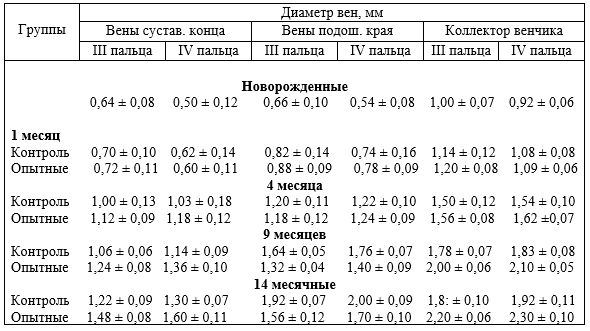
При ежедневном ДПД на расстояние 3,5–4,0 км/час венозные сети становились плотнее, сохраняются слои, диаметр вен увеличивается в среднем 1,16–1,32 раза, при этом вены становятся крупнее, ячеи сети, снимается отёчность в области акроподия (рис. 24).
25. Вены подошвенной поверхности копытца
Отток крови из подошвенной поверхности копытца животных осуществляется
по аксиальным и абаксиальным венам, диаметром 0,1 – 0,25 мм, что значительно ниже по диаметру вен париетальной поверхности (0,1- 0,5 мм) и межкопытцевой
(0,1-0,3 мм).
Рис. 24. Густые венозные сети основы кожи париетальной
поверхности копытца, гипердинамия. Опыт, бык 14 месяцев.
В точке выхода вен эмиссарий из III фаланги, располагаются двухстворчатые клапаны, направляющие кровоток в венозные сети основы кожи подошвенной поверхности копытца. Из этой сети выходят специальные вены III и IV пальца. Они располагаются в сосудистом жёлобе плантарной поверхности копытца. Желоб исключает передавливание вен челночной и копытцевой костями.
В образовавшуюся специальную вену открываются вены мякиша и другие приточные венозные ветви. Число эмиссарий и суммарный диаметр вен подошвенной поверхности копытца, изменяется с возрастом и зависит от двигательной активности животного. Изменения наступают обычно с возраста телят четырёх месяцев. Так, при гиподинамии (контрольная группа) число истоков в возрасте девяти месяцев достигает 3–5, 14 месяцев 4–5. В этой зоне копытцевой кости, молодняк получавший ежедневно ДПД 3,5–4,0 км/час, в возрасте девяти месяцев истоков было 4–6, а в 14 месяцев доходило до семи.
Суммарный диаметр вен подошвенной поверхности копытца новорожденных телят достигал 0,7 мм, У телят контрольной группы в месячном возрасте был 0,83 мм, в четырёхмесячном – 1,23 мм, в девяти – 1,5 мм, а в 14-месячном – 1,75
мм. В опытной группе, получавшей ДПД, соответственно, был выше в 1,1; 11,17;
1,27 и 1,36 раза. Но суммарный диаметр вен париетальной (спинковой) поверхности был больше, чем подошвенной (табл.49).
49. Возрастная динамика вен копытцевой кости III пальца тазовой конечности бычков
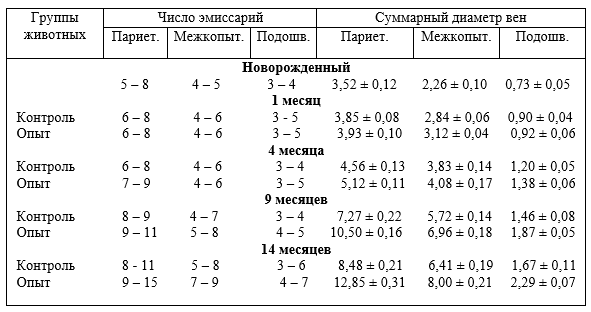
Из таблицы видно, что число выпускных отверстий и диаметр вен копытцевой кости III пальца начинает увеличиваться с возраста телят четырёх месяцев. Особенно интенсивно возрастает диаметр вен париетальной, несколько ниже - аксиальной и слабо – подошвенной поверхности. В целом диаметр вен опытной группы превосходит калибр вен контрольной, начиная с четырёхмесячного возраста и до 14 месяцев в 1,12 – 1,52 раза.
Между морфометрическими показателями экстраорганных вен копытца III и IV пальца достоверных различий не обнаружено, но замечено, что число истоков, суммарный диаметр вен копытца IV пальца был выше, чем Ш пальца, особенно это видно в опытной группе (табл. 50).
27. Венозный коллектор венчика копытца
Коллектор располагается в подкожном слое венчика и выполняет функцию гемоамортизатора и биологического устройства, откачивающего кровь из копытца. Между роговой капсулой и третьей фалангой пальцев мышц нет, вены копытца все тонкостенные и обеспечить отток крови из пальца не могут. Поэтому, механизм оттока крови из коллектора происходит поэтапно, следующим образом:
Первый этап – в случае жёсткого грунта крупный рогатый скот опирается о почву подошвенным краем париетальной поверхности копытца и дополнительно
подошвенной поверхностью – в случае мягкого грунта. В копытце жвачных отсутствуют заворотные углы и стрелка, имеющаяся у лошади. Сзади между краями аксиальной и абаксиальной поверхности роговой стенки копытца располагается мякиш, который глубоко вдаётся в зацепную часть между основой кожи и подошвенной поверхностью копытца. При опоре конечности о почву копытцевая кость вдавливается в роговую капсулу, мякиши III и IV пальцев, образно раздавливаясь, разводят задние края париетальной стенки копытца, увеличивая объём их венозной сети основы кожи (см.гл.25). В сеть устремляется кровь из основы кожи подошвенной поверхности. Назад кровь течь не может, так как в соустьях притоковых вен имеются клапаны.
50. Возрастная динамика вен копытцевой кости IV пальца тазовой конечности
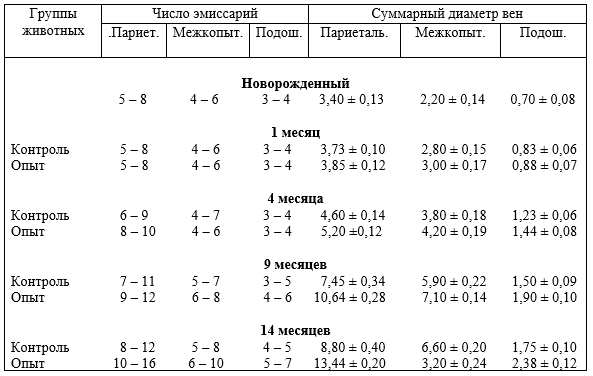
Второй этап. При поднятии конечности задние края аксиальной и абаксиальной поверхности париетальной стенки капсулы копытца под действием обратной упругости рога сходятся. Вены основы кожи сдавливаются и кровь по венам эмиссариям устремляется в коллектор венчика копытца. В обратном направлении течь она не может, так как в соустьях их имеются клапаны.
Третий этап. При первоначальной опоре о грунт торцовое давление роговой капсулы передаётся на основу и подкожный слой венчика копытца. В подкожном
слое располагается коллектор венчика, из которого кровь выдавливается в общие
дорсальные, пальмарные (плантарные) пальцевые вены. Обратно кровь поступать не может, так как в соустьях притоковых вен располагаются двухстворчатые клапаны. Таким образом венозный коллектор венчика выполняет как гемоамортизационную функцию, так и устройства подающего кровь проксимально из копытца в поверхностную и глубокую венозные магистрали конечностей.
Из соединения правого и левого коллекторов на дорсальной поверхности кисти (стопы) проксимально выходит общая дорсальная III и IV пальца вена. На ладонной поверхности коллектор венчика III и IV пальца объединяет ладонный или пальмарный (плантарный или подошвенный) поверхностный анастомоз, из которого выходит проксимально общая пальмарная (плантарная) III и IV пальца вена (рис.25).
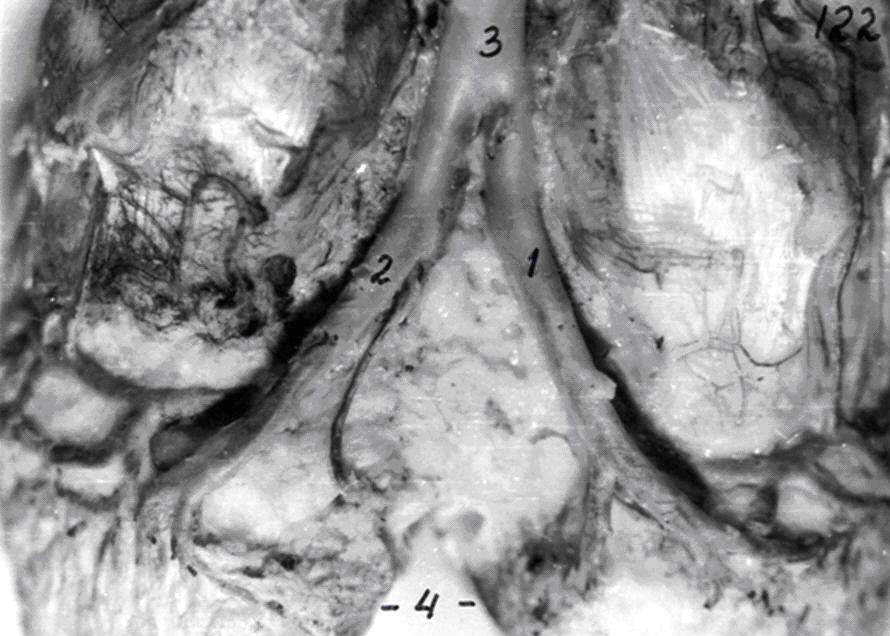
Рис. 25. Слияние венозных коллекторов III и IV пальцев.
1 - осевая собственная дорсальная вена IV пальца; 2 – осевая собственная
дорсальная вена III пальца; 3 – общая дорсальная вена III и IV пальца;
4 – свод межкопытцевой щели.
Диаметр вен коллектора в месячном возрасте телят, содержащихся в клетках (при гиподинамии), равен 1,08 мм, в четыре месяца – 1,54 мм, в девять – 1,83 мм и 14 месяцев – 1,92 мм. У телят, получавших ДПД, в месячном возрасте почти не отличался, но начиная с четырёх месяцев и выше, соответственно был больше в 105; 1,15 и 1,2 раза. Как видно разница существенная.
28. Общая дорсальная вена III и IV пальца
Общая дорсальная вена выходит из слияния венозных коллекторов III и IV копытцев. Поднимается проксимально в плотном фасциальном влагалище, в
дорсальной продольной борозде, под кожей. В области дистального канала пясти (плюсны) участвует в образовании глубокой пальмарной (плантарной) дуги (рис.26). Затем смещается на дорсолатеральную поверхность пясти (плюсны) и открывается в поверхностную венозную магистраль грудной конечности (см.гл.21) или в ветви латеральной вены сафены – тазовой конечности (см. гл. 23).
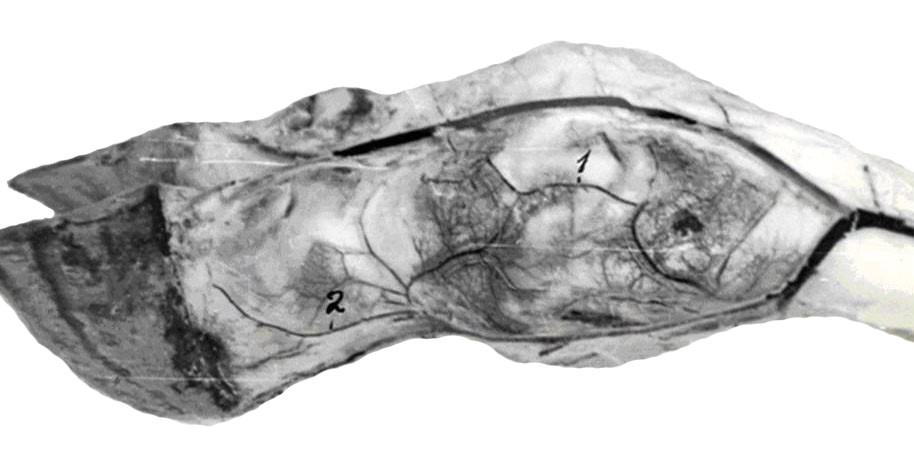
Рис. 26. Притоковые ветви общей дорсальной вены III и IV пальцев.
Диаметр вены изменяется не только с возрастом молодняка, но и под действием дополнительного принудительного движения (ДПД). Так, новорожденных телят диаметр вены равен 1,9 мм, в месячном возрасте контрольной группы – 2,1 мм, в четыре месяца – 3,76 мм, в девять – 4,08 мм и в 14 месяцев – 4,3 мм.
В опытной группе, получавшей ДПД, в месячном возрасте превосходила контрольную - в 1,01 раза, в остальных соответственно – в 1,04, 1,1 и 1,2 раза. Разница не существенная, но, согласитесь, имеется.
Клапаны располагаются в соустье притоковых вен и при открытии в поверхностные венозные магистрали конечностей, чаще двухстворчатые. Число клапанов внутри ствола вены колеблется от двух до четырёх. При этом длина вены достигает в пределах 15,2 см. Клапанный индекс составляет 5, 0 – 7,6 см и иногда клапанный индекс в вене составляет 8–14 см.
29. Вены латеральной и ладонной (подошвенной) поверхности копытца
В целом вены пальцев можно разделить на дорсальную, латеральную, пальмарнo-ладоннную (плантарную-подошвенную) системы, т.к. в связи терминологией и большим числом анастомозов, венечных дуг, тяжело в этом разобраться. Считаем, что систематика в этом вопросе поможет читателю, специалисту.
Дорсальная, париетальная, подошвенная система вен была рассмотрена в 24 – 28 главах. Латеральная (боковая) система вен представлена подкожными абаксиальными венами III и IV пальцев. Обе подкожные вены располагаются в подкожном слое второй фаланги. Основной ствол их начинается притоковыми кожными ветвями нижней латеропальмарной (латероплантарной) роверхности второй фаланги третьего и четвертого пальца. В него открываются несколько ветвей из коллекторов венчиков копытцев, мякишей. Вены, поднимаясь проксимально, увеличиваются в диаметре (рис.27).
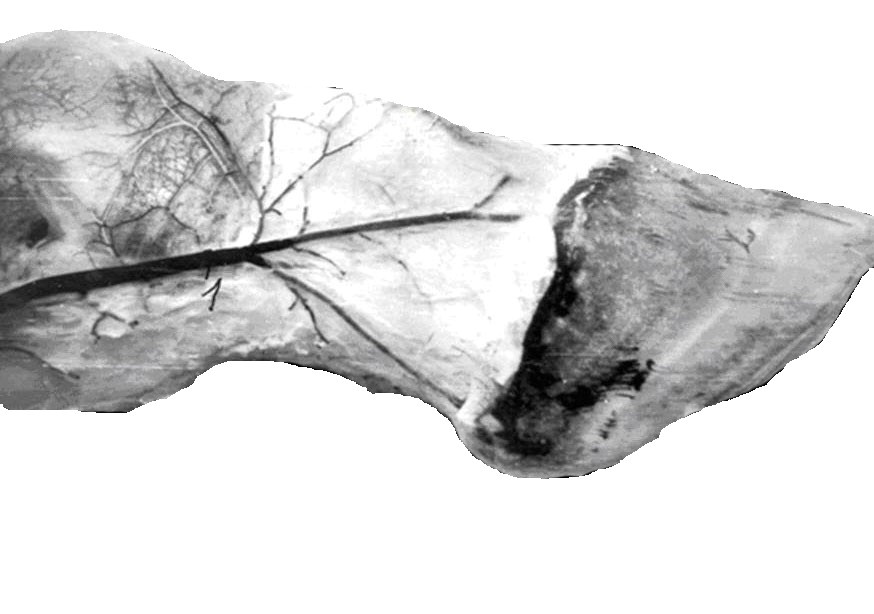
Рис.27. Подкожная абаксальная (1) вена III пальца.
Бык 14 месяцев.
Стволы вен располагаются в фасциальных влагалищах, по бокам сухожилий поверхностного и глубокого пальцевых сгибателей. На середине тела второй фаланги они анастомозируют (соединяются) между собой и с аксильными глубокими венами III и IV пальца, в соустьях их находятся двухстворчатые клапаны и через межпальцевую щель вступают в анастомоз с общей дорсальной веной III и IV пальца.
По ходу подкожные абаксиальные вены III и IV пальца принимают притоковые ветви сетей суставных капсул и рудиментов пальцев. Выше головок III и IV пястных (плюсневых) костей вены под третьей межкостной мышцей вступают в анастомоз с глубокой пястной (плюсневой) дугой, а на задней конечности дополнительно с латеральной ветвью наружной вены сафены. Из этого следует что кровь может оттекать как в глубокую, так и в поверхностную венозные магистрали конечностей. Всего по ходу вена принимает до 16 притоковых ветвей, диаметром 0,2 – 1,4 мм. Подкожная абаксиальная вена IV пальца новорожденного достигает 1,8 мм, в месячном возрасте – 1,9 мм, в четырёх – 3,1 мм, в девяти месячном – 4,1 мм и 14 – месячном – 4,3 мм. У телят, получавших ДПД, калибр вен был выше, соответственно – в 1,01; 1,05; 1,11 и 1,1 раза. Разница не существенная для вен, но все же имеется (рис. 28).
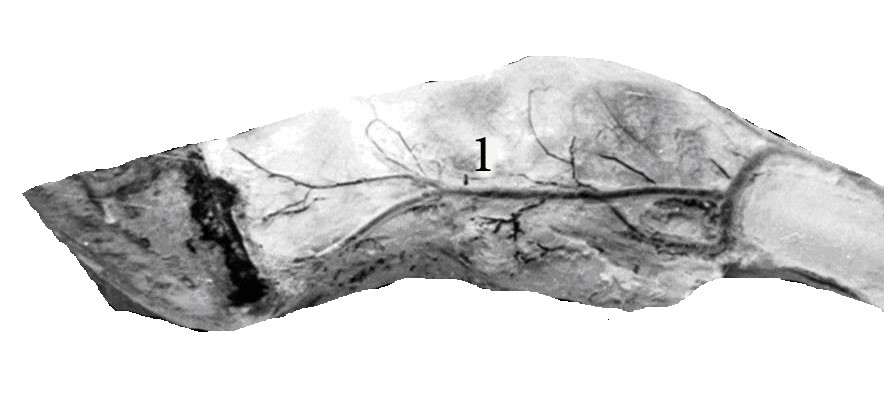
Рис. 28. Подкожная абаксиальная (1) вена IV пальца, бык 14 месяцев.
В просвете вен и в соустьях притоковых имеются развитые двухстворчатые клапаны. Что касается длины вен. Она целиком попадает в зависимость от развития и роста тела и конечностей в длину и высоту. По этому, чем полноценнее будет рацион питания, тем интенсивнее будет рост организма и вен.
Глубокая плантарная (пальмарная) система вен образуется слиянием осевых и неосевых собственных вен III и IV пальца. Они, в свою очередь, сливаясь, формируют общие плантарные (пальмарные) вены III и IV пальца. В общие вены третьего и четвёртого пальца открываются в вены, выносящие кровь из второй фаланги, мякишей.
Общие подошвенные (ладонные) вены III и IV пальца, сливаясь, формируют в начале межпальцевую, а затем она, поднимаясь, проксимально переходит в общую пальцевую вену. Таков запутанный номенклатурный расклад вен.
И так, аксиальные (осевые) и абаксиальные (неосевые) вены III и IV пальца (каждая кость имеет одну осевую и неосевую вену) короткие, тонкие, выходят из одноименных отверстий копытцевых костей, сливаясь, образуют осевые и неосевые собственные плантарные (пальмарные) вены III и IV пальца. В начале вены располагаются сбоку сухожилий глубокого сгибателя пальцев, по ходу принимают до шести притоковых вен из мякишей, коллекторов венчика копытец,
основы кожи париетальной поверхности копытцев, суставов второй и третьей фаланги и переходят в общие плантарные ( пальмарные) вены III и IV пальца. По ходу в них открываются венозные ветви второй и третьей фаланги и на уровне нижнего края межголовчатой вырезки пястья (плюсны) объединяются в межпальцевую вену.
Дорсальная (спинковая) и плантарная (пальмарная) межпальцевые вены в этом месте соединяются между собой венозной дугой. Из соединения проксимально поднимается общая пальцевая вена, которая в области дистального прободающего канала плюсны (пясти) участвует в образовании глубокой плантарной (пальмарной) дуги.
Общая плантарная пальцевая вена располагается поверхностно под кожей, в жёлобе, образованном сухожилиями поверхностного и глубокого сгибателей пальцев, проходит между сезамовидными костями на плантарную поверхность плюсны (пястья), погружается под третью межкостную мышцу. В этом месте получает анастомоз через дистальный канал плюсны от общей дорсальной пальцевой вены и сюда вливаются подкожные абаксиальные вены III и IV пальца. В итоге образуется глубокая планатарная (пальмарная) дуга, из которой проксимально выходит глубокая венозная магистраль тазовой конечности (рис.29, 30).
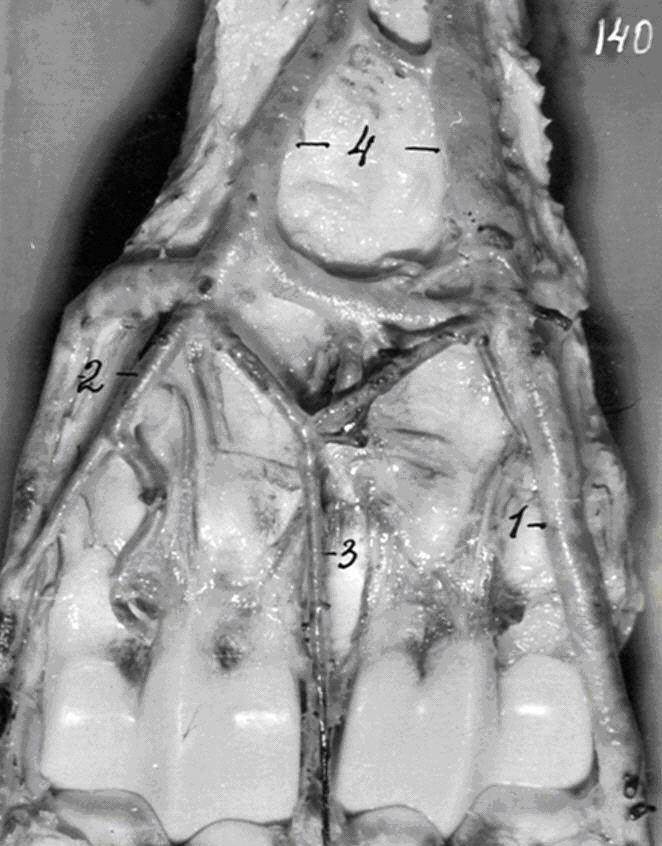
Рис.29. Глубокая плантарная (пальмарная) дуга.
1, 2 – подкожная абаксиальная вена III и IV пальца; 3 – общая
пальцевая вена; 4 – глубокие плюсневые вены. Бык 14 месяцев.
Как формируется поверхностная венозная магистраль грудной конечности описано в главе 21. Тазовой конечности поверхностная венозная магистраль представлена латеральной и медиальной венами сафена. Латеральная вена сафена выходит краниальной ветвью из поверхностной дорсальной дуги, образованной слиянием общих дорсальных вен III и IV пальцев. Поверхностная дорсальная дуга соединяется через дистальный плюсневый канал с плантарной глубокой дугой. Вторая, каудальная ветвь латеральной вены сафены выходит из анастомоза задней ветви медиальной вены сафены. Из этого следует, что из акроподия кровь может оттекать, как через латеральную, так и медиальную вены сафена (см. гл. 23) и через глубокую венозную магистраль.
Диаметр общей пальцевой вены, молодняка содержащегося в условиях гиподинамии, в месячном возрасте составил 1, 45 мм, в четырёхмесячном – 1,74 мм, в девятимесячном – 2,06 мм, в 14 месяцев был равен – 2,28 мм. Получавших ДПД, был выше, соответственно – в 1,01; 1,03; 1,05 и 1,06 раза. Из данного анализа следует, что у животных получавших ДПД, диаметр вен был выше, чем молодняка клеточного содержания.
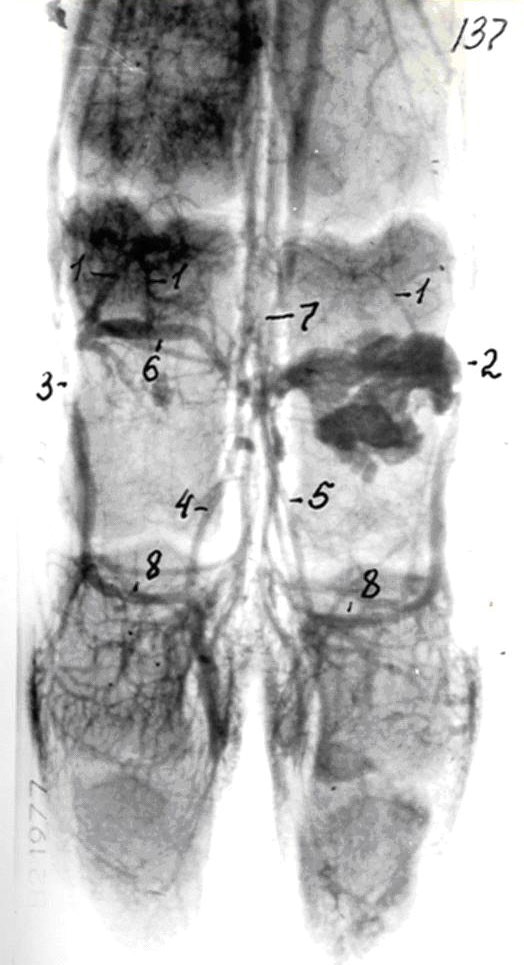
Рис. 30. Рентгенограмма вен плантарной поверхности акроподия.
1 – вены проксимального конца первой фаланги;
2, 3 – абаксиальные поверхностные вены III и IV пальца; 4,5 –
аксиальные плантарные вены III и IV пальца; 6 – вена второй
фаланги; 7 – межпальцевая вена: 8 – венозный коллектор венчика.
Клапанный индекс общей пальцевой вены молодняка клеточного содержания был равен 0,32–0,43 см, а получавших ДПД 0,33–0,46. Клапаны общей пальцевой вены чаще располагаются в устьях притоковых вен, реже в просвете вены. Но в 80% случаев обнаружено, что нижний устьевой клапан открывается кармашками вниз, а не вверх. Из этого следует, что отток крови из вены может происходить не проксимально - в глубокие плюсневые вены, а через плантарную глубокую дугу – в поверхностную венозную магистраль. По этому, число клапанов в её просвете варьирует от одного до четырёх. В вене контрольной и опытной групп, независимо от степени двигательной активности, разницы между клапанными индексами не обнаружено. Однако замечено, что в венах IV пальца с возраста четырёх месяцев наблюдается тенденция к увеличению морфометрических показателей (длины, диаметра) по сравнению с таковыми венами Ш пальца, которая больше проявляется у молодняка опытной группы.
Вопросы для контроля
- Какими фалангами представлен скелет пальцев?
- Строение фаланг, к какому типу костей их можно отнести?
- Строение копытцевой кости?
- Что за отверстия эмиссарии и какое их назначение?
- Какие вены выносят кровь из копытцевых костей?
- Какие вены выносят кровь от пальцев?
- Какие сети в копытце формируют вены?
- Как изменяется венозная сеть спинковой (дорсальной) поверхности копытца при гиподинамии и при ДПД?
- Что представляет венозный коллектор копытца, где располагается и его функция?
- Механизм оттока крови из венозного коллектора копытца?
- Как формируется общая дорсальная вена III и IV пальцев?
30. Динамические нагрузки и сосуды пальцев
Известно, что в промышленных комплексах крупный рогатый скот круглый
год содержится в помещениях на решетчатых или бетонных и редко на деревянных полах. Считается, чем меньше скот движется, тем больше питательных веществ усваивается организмом, тем больше привесы. Частично это так. Но опыт промышленных комплексов показывает, что на решётчатых, бетонных полах чаще возникают различного рода ушибы, растяжения сухожилий, разрывы межпальцевых связок, тендовагиниты, артриты, некробак-
териоз и т.д. Вследствие чего происходит досрочная выбраковка животных по причинам травматизма копытец, появления трещин, язв, абсцессов, панариций и др. (П.И.Баканов, 1978).
Морфология каймы, венчика и полностью копытца дана в главах 24 и 25, и разъяснена их особенность строения. Рог абаксиальной, зацепной поверхности копытца толще, чем аксиальной. В венчике копытца абаксиальной поверхности находится больше крупных артерий и вен.
У молодняка первой (контрольной) группы, при клеточном содержании на решётчатых полах, выявлены следующие изменения сосудов. При гиподинамии происходит, прежде всего, частичное запустевание и облитерация микроциркуляторного русла сосочкового слоя, основы кожи каймы и венчика копытца. Артерии дермы и субдермы венчика меняли толщину стенок и просвет сосуда. Стенки артерий по толщине превосходят диаметр почти в три раза, т.е. просвет артерии составлял одну треть от диаметра, а местами наблюдалась облитерация артерий, но структура стенок вен, при этом, не менялась, что может привести к остеопорозу, некрозу костной ткани (рис. 31).
Сама форма артерий и вен подвергалась деформации. Артерии по периметру становятся округлыми, стенки толстые, как бы отёчны, интима представлена крупной складчатостью. Поэтому поступление питательных веществ из крови в костную ткань становится проблематичной. Вены сплющены, в поперечном сечении овально вытянутые (рис. 32).
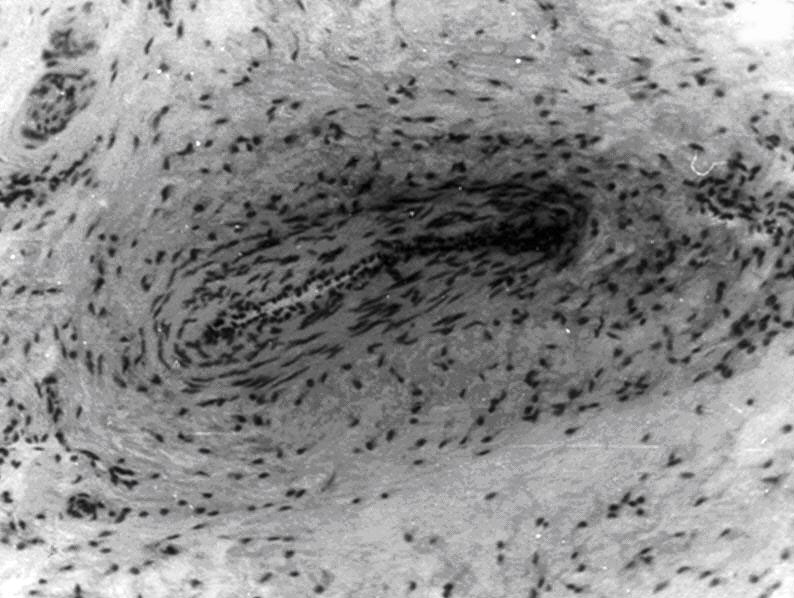
Рис.31. Идёт облитерация артерии.
Контроль, бык 9 месяцев.
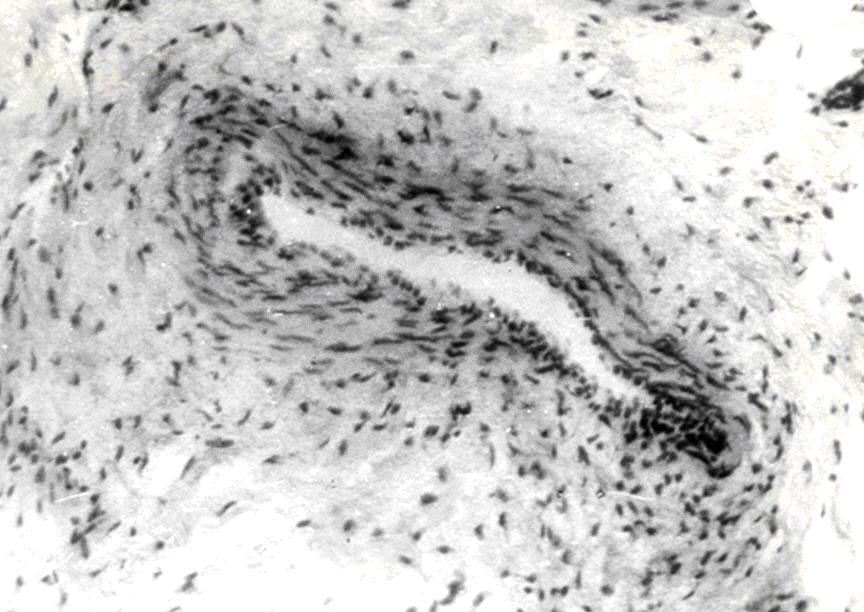
Рис. 32. Поперечный срез вены второй фаланги. Контроль,
бык четыре месяца
Меняется и микроциркуляторное русло, часть артериол и прекапилляров запустевает, а часть остаётся разреженной. Это приводит к нарушению крово-обращения в данной точке и сосочкового слоя основы кожи каймы и венчика копытца и, в конечном счёте, к нарушению образования глазури и трубчатого слоя роговой капсулы копытца (см.гл. 24, 25).
«Полями», роговая стенка копытца имела нормальную толщину стенки (4-7 мм), а местами – тонкую (3–5 мм). По этому (см. рис. 23), нормальное кровоснабжение росткового слоя эпидермиса, основы кожи венчика копытца имеет большое значение для накопления кератина и образования роговой капсулы копытца. В «поле» с тонкой стенкой роговой капсулы, в сосочковом слое основы кожи сети имели разреженные петли, что приводит к недостаточному накоплению клетками росткового слоя кератина, отсюда нарушение образования рога, появляется хромота, трещины, рыхлость рога копытца, куда проникает анаэробная инфекция, вызывает воспаления, появляются язвы, копытная гниль и т.д. Отсюда понижается экономическая эффективность, падёж и другие убытки.
Стенка экстраорганных вен состоит из интимы, меди и экстерны (рис.33). В просвете костных каналов стенки вен тонкие и их медия представлена единичными гладкими мышечными клетками. По мере удаления вены от кости количество гладких мышечных клеток и эластических волокон в средней оболочке их стенки увеличивается, а в устье с магистральными венами они образуют сфинктеры (см. рис.10 и 11).
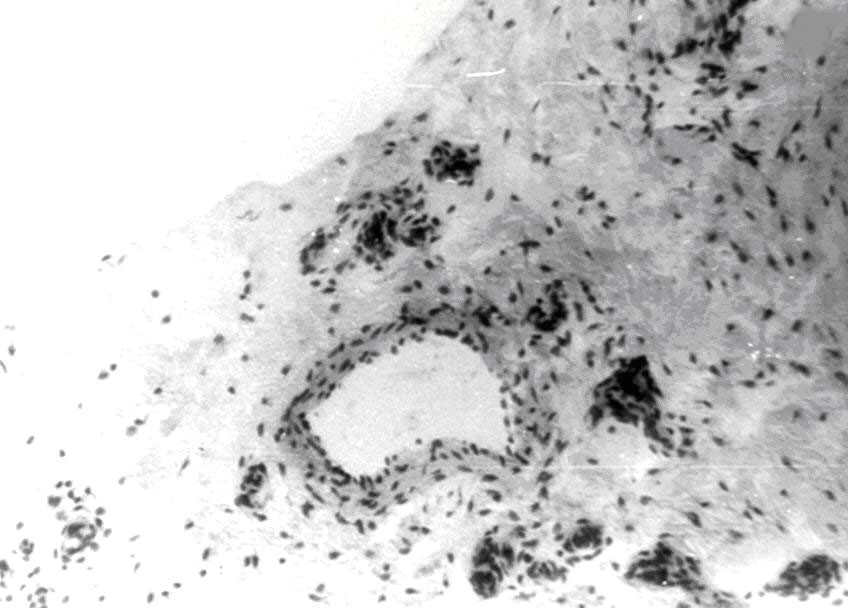
Рис. 33. Срез вены плантарной поверхности второй
фаланги. Контроль, бык четыре месяца.
Толщина стенки вен дистального конца второй фаланги при выходе из кости новорожденного телёнка достигает 6,60 мкм, в месячном возрасте клеточного содержании уже - 7,75 мкм, в четырёхмесячном возрасте – 9,16 мкм, девяти – 15,62 мкм и в 14 месяцев – 16,30 мкм (табл.51).
51. Динамика толщины стенок вен второй фаланги III пальца тазовой конечности
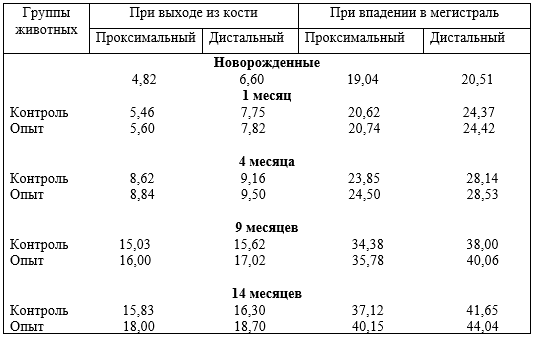
Молодняк, получавший ДПД (опытная группа) толщина стенки вен увеличивалась до четырёхмесячного возраста, в пределах допустимой ошибки, но начиная с девяти и до 14 месяцев, превышала контрольную группу, соответственно в 1,09 и 1,15 раза, что существенно, а в абсолютных величинах
было 2,54–3,2 мм. Для сравнения стенка капилляра имеет толщину в пределах 1,8–2,2 мкм.
Все вены эмиссарии верхнего конца второй фаланги имели диаметр и толщину стенок значительно тоньше, чем нижнего. Они выходят из отверстий через желоб, который предохраняет их от механических пережатий сухожилиями, связками. Поднимаясь проксимально, по ходу принимают приточные венозные ветви из надкостницы фаланги, капсул суставов и вливаются в плантарный анастомоз, соединяющий венозные коллекторы III и IV пальца, а часть - в поверхностные аксиальные и абаксиальные кожные вены.
Клапанный индекс вен второй фаланги равен 1,8–4,2 мм и клапаны представлены двумя створками. В целом, между морфологическими показателями экстраорганных вен второй фаланги III и IV пальца не обнаружено. Но установлено, что с возрастом морфометрические показатели вен
IV пальца явно имели превосходство над венами фаланги III пальца. Эта тенденция более выражена у животных опытной группы, получавшей дозированное принудительное движение.
Вопросы для контроля
- Что происходит с артериями копытца при гиподинамии?
- Что происходит с венами копытца при гиподинамии?
- Как изменяется просвет артерий и вен при гиподинамии и при гипердинамии?
- К каким заболеваниям копытц может привести гиподинамия?
31. Влияние опоры и угла наклона копытца на вены акроподия
Из предыдущей главы видно, что на при гипо и гипердинамии на гемодинамику акроподия и на возникновение заболеваний непосредственно влияет роговое копытце. В связи с чем, возник вопрос – изменяется или нет характер опоры и угол наклона копытца при различной динамической нагрузке. Как отражается это на гемодинамике копытца. Для уточнения были подобраны контрольная и опытная группы по принципу аналогов. Контрольная группа содержалась в клетках, а опытная получала до 3,5 - 4 км/час ДПД.
Исходными данными были площадь опоры и угол постановки копытца новорожденных телят. Площадь опоры IV пальца новорожденного телёнка достигала 6,51 см2 . У телят контрольной группы в месячном возрасте она
составила 8,63 см2 , в четыре месяца - 12,08 см2 , в девять – 20,16 см2 , а в четырнадцать месяцев – 22,55 см2 . Площадь опоры опытной группы, получав-
шей ДПД, превышала контрольной, соответственно в 1,0; 1,04; 1,21 и 1,39 раза
(табл.52).
52. Площадь опоры копытец тазовой конечности
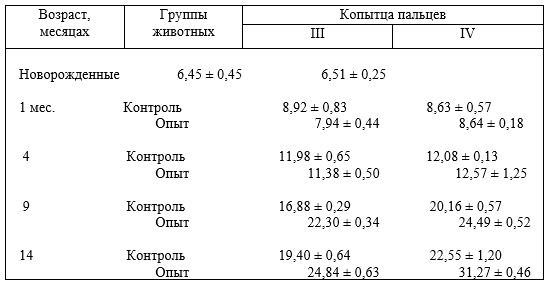
Из таблицы видно, что площадь опоры копытца IV пальца достоверно больше площади опоры копытца III пальца как в опытной, так и в контрольной группах. Так при гиподинамии площадь опоры копытца IV пальца была больше Ш пальца в девять месяцев на 3,3 см2 , а в 14 месяцев – 3,15 см2 , а получавших ДПД, соответственно больше в 1,01 и 2,04 раза. Разница существенная.
Угол наклона IV пальца новорожденных телят составил 50,3о. Молодняка контрольной группы в месячном возрасте достиг 48,4о, в четыре – 52,6о, в девять – 55,6о, а в 14 месяцев – 59,3о. В опытной группе, получавшей ДПД, был ниже, чем в контрольной, соответственно, на 0,9о; 1,4о; 3,4о; и 4,6о. Увеличение угла наклона пальцев в контрольной группе, содержащихся в условиях гиподинамии, приводит к образованию «торцового копытца» и к уменьшению площади опоры. Это значит, что связки обладают слабой эластичностью, плохо растяжимые, чаще
подвергаются растяжению, микротравмам, а движения в суставах ограничены, тугие. В итоге - это, как правило, приводит к травмам суставов.
Исследование показало, что угол наклона внутреннего (III) пальца контрольной и опытной групп был больше, чем наружного, но в контрольной группе угол наклона пальцев меньше, чем - в опытной. Торцовость копытец обычно наблюдается у животных, обитающих на жёстком каменистом грунте, на
цементных, решётчатых и щелевых полах. Об этом имеются пояснения в научной литературе (В.Б.Борисевич, 1983; В. Раднаев, 1992). Авторы отме-
мечают, что копытца различных пород крупного рогатого скота имеют неодина-
ковый угол наклона, у симменталов - он выше, а чёрно-пёстрой породы - ниже. Причём медиальное копытце крупного рогатого скота грудной конечности, по сравнению с латеральным, имеет более тупой угол наклона в зацепе, что обусловлено более выраженной статистической функцией третьего пальца. На тазовой конечности копытце имеет остроугольную форму, что совпадает с мнением ряда авторов (К.А.Васильев, 1991) о превалировании этого пальца в работе тазовой конечности. Наши данные приведены в таблице 53.
При «торцовым копытце» париетальная и подошвенная поверхности роговой капсулы копытца сдавливают венозные сети основы кожи, затрудняя отток крови
в венозный коллектор венчика. В связи с этим, отток крови из копытца происходит не в проксимальном направлении, а из терминальной дуги в пальмарном (плантарном) направлении, что приводит к ослаблению гемоамортизационных функций венозного коллектора и возможно к развитию гематом, отёков и других осложнений в области венчика копытца.
53. Угол наклона копытец тазовой конечности бычков
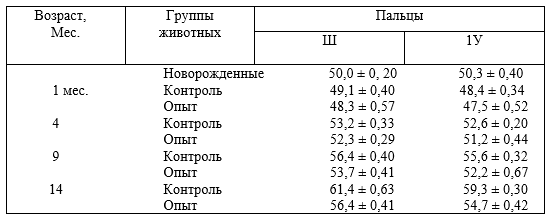
Вопросы для контроля
- Влияет или нет угол наклона на структуру вен пальцев?
- У какого пальца торцовость выше?
32. Прочность костей акроподия
Венозное русло костей пальцев тесно связано с функциональной нагрузкой и структурой костной ткани. Оказывает влияние на её биофизические свойства,
которые отражают функциональное состояние костной ткани, связанных в одну структуру. Исследование прочности костей пальцев показало, что они имеют не только возрастные особенности, но и зависят от степени двигательной активности животных и расположения в звене.
Прочность первой фаланги IV пальца новорожденных телят выдерживает давление на грань 263,2 кг/см2. У телят, содержавшихся в клетках (гиподинамия), в месячном возрасте выдерживали давление на грань 308,4 кг/см2, четырёхмесячном – 415,6 кг/см2, девятимесячном – 630,3 кг/см2 и в 14 месяцев – 807,1 кг/ см2. У телят, получавших ДПД, прочность костей в месячном возрасте была выше в 1,01 раза, а в остальных, соответственно – 1,06; 1,17 и 1,13 раза. Из этого следует, что прочность первой фаланги в обеих группах молодняка до четырёх месяцев почти не менялась и существенно возрастает у животных в девять и 14 месяцев, получавших ДПД.
Прочность второй фаланги IV пальца новорожденного телёнка на излом несколько ниже, чем первой и достигает 234,6 кг/см2. У телят месячного возраста клеточного содержания она достигала 236,2 кг/см2, в четыре месяца – 390,5 кг/см2, в девять – 550,3 кг/см2 и в 14 месяцев – 602,2 кг/см2. У телят, получавших ДПД, было, соответственно, выше – в 1,02; 1,05; 1,17 и 1,17 раза. Эти данные свидетельствуют, что прочность второй фаланги молодняка наиболее повышается с четырёх до девяти месяцев, получавшего ДПД, и до 14 месяцев сохраняется на этом уровне. На что указывает коэффициент достоверности опытной группы.
Прочность копытцевой кости (III фаланги) IV пальца новорожденного телёнка выдерживает давление на грань 184,8 кг/см2, месячном возрасте телят клеточного содержания – 189,6 кг/см2, в четыре – 216,2 ег/см2, девять – 340,4 кг/см2 и в 14 месяцев – 408,3 кг/ см2. У телят, получавших ежедневно дозированное принудительное движение, кость которых выдерживала давление на разрушение, соответственно, коэффициент был выше - 1,01; 1,08; 1,21 и 1,16. Это большая разница. Из анализа следует, что прочность кости интенсивнее возрастает у молодняка, получавшего ДПД, с четырёх до 14 месяцев, но наиболее активно возрастает в девять месяцев.
Из сравнения видно, что наиболее прочным в контрольной и опытной группах является первый фаланг, затем – второй и третий (табл.54).
54. Прочность фаланг пальцев III и IV пальцев тазовой конечности (кг/см2)
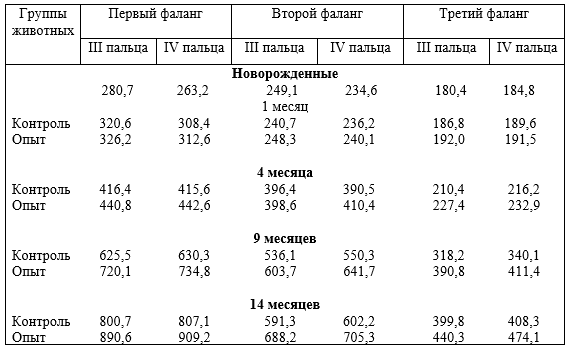
Исследования показали, что суммарная величина диаметра вен на выходе из отверстий эмиссарий костей пальцев, их масса и прочность у животных с возрастом увеличивается неодинаково и зависит от степени двигательной активности, и расположения костей в звене акроподия. Так, суммарный диаметр вен путовой и венечной кости в опытной группе, получавших ДПД, наиболее интенсивно увеличивается с месячного до четырёхмесячного возраста, в контрольной группе значительно меньше. Предполагаем, что в период с одного до четырёх месяцев, когда масса тела телят незначительная, идёт интенсивная подготовка сосудистого русла для дальнейшего развития кости и становления её биофизических свойств как опорной конструкции. Что подтверждается ростом массы и прочности фаланг пальцев с четырёх месяцев до девяти (табл. 55).
Это же происходит и с копытцевой костью, но следует обратить внимание на то, что суммарная величина диаметра вен копытной кости во всех возрастных периодах была больше, чем первого и второго фаланг. Значение этого показателя для молодняка, получавшего ДПД, с месячного возраста достоверно больше, чем у животных клеточного содержания, что свидетельствует об отрицательном действии гиподинамии на морфофункциональное состояние сосудистого русла
пальцев.
55. Суммарный диаметр вен костей акроподия тазовой конечности бычков (мкм)
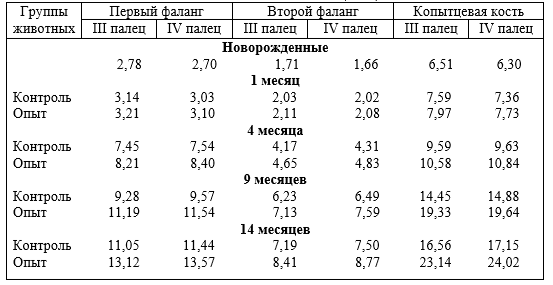
Подобные данные были получены при исследовании компактного костного вещества костей стилоподия и зейгоподия северного оленя и крупного рогатого скота (см.гл.15 и 16). При гиподинамии у крупного рогатого скота компактное костное вещество становится толще, рыхлее, остеоны становятся в диаметре больше, разреженно расположенные, гаверсовы и фолькмановы каналы по диаметру тоньше, а при ДПД, наоборот, компактное костное вещество становится тоньше, плотнее, остеоны меньше по диаметру и увеличивается количество их в поле микроскопа, все каналы шире. Особенно это заметно в костях северного оленя, ведущего полудикий образ жизни.
Вопросы для контроля
- Повышает или нет прочность костей пальцев ДПД?
- Какой фаланг наиболее прочный на излом?
- Какая кость имеет более плотное компактное вещество, получавших ДПД или нет?
33. Возрастные изменения массы костей пальцев задней конечности в зависимости от ДПД
Скелет выполняет ряд важных функций в организме. Мы не будем останавливаться на известных функциях: защиты, опоры, рычагов движения, кроветворения (Х.Б.Баймишев, Б.П.Шевченко, М.С.Сеитов, 2009). Но на некоторых следует остановиться. Скелет, в целом, является минеральным депо в
организме, а отсюда гемопоэтическая, электролитическая, обменная и другие функции. Большое значение в организме имеют такие минералы как: фосфор, калий, кальций, натрий, магний, хром, ванадий и др. Особенно велико значение кальция для нервной и опорной систем. Что важно для понимания механизма развития у беременных животных родильного пареза, остеомаляции, остеопоро-
за, остеохондроза, у молодняка рахита и т.д.
Вены костей тесно связаны как с костной тканью, красным и жёлтым мозгом, так и с нервной системой, органами внутренней секреции. Поэтому они тесно контактируют с костной тканью, её механическими, биохимическими процессами, которые протекают в ней и оказывают как положительное, так и отрицательное влияние на костную ткань.
Проведенные исследования позволили установить, что рост массы костей пальцев тазовой конечности бычков, увеличивается в соответствии с общими биологическими законами развития тканей. Функция воздействует на материю, материя – на функцию. При понижении функции – атрофируется материя на степень уменьшения функции и, наоборот – при повышении функции, масса материи гипертрофируется на степень повышения функции. Поэтому масса костей пальцев попадает в зависимость от ДПД и положения в звене пальца.
Масса первой фаланги IV пальца тазовой конечности новорожденных телят достигает 14,6 г. В возрасте одного месяца клеточного содержания составляет - 21,5 г, в четыре – 30,9 г, девять – 68,7 г и в 14 месяцев – 76,8 г. Масса этой кости телят, получавших ДПД, была больше, соответственно, с четырёх месяцев – в
1,09; 1,04 и в 1,1 раза.
Масса первой фаланги IV пальца задней конечности до возраста четырёх месяцев повышается одинаково как в контрольной, так и опытной группах. Но в старших возрастах этой группы эта разница стала чётко возрастать и в 14 месяцев была выше в 1,1 раза, чем в контрольной..
Масса второй фаланги IV пальца новорожденных телят равна 11,7 г. В месячном возрасте телят клеточного содержания составила 12,8 г, в четыре месяца – 19,1 г, девять – 41,1 г и в 14 месяцев – 46,0 г. У телят ежедневно получавших ДПД на расстояние 3,5 – 4,0 км/час была, соответственно, больше –
в 1,08; 1,11; 1,07 и 1,07 раза. Таким образом, масса второй фаланги приростает по закономерности первой, но разница в приросте её уже заметна с месячного возраста (табл.56).
56. Динамика роста массы костей пальцев бычков задней конечности (г)
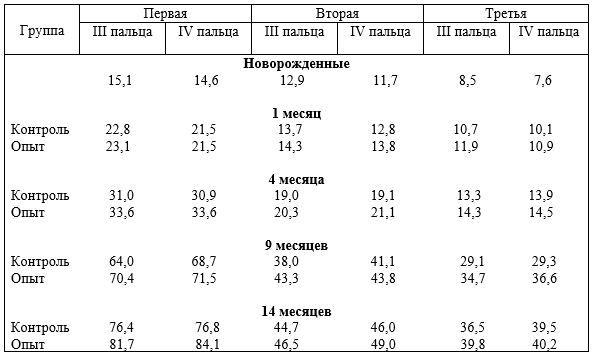
Масса третьей фаланги (копытцевой кости) новорожденных телят составила 7,6 г, в месячном возрасте телят клеточного содержания - 10,1 г, в четыре месяца – 13,9 г, девять – 29,3 г и в 14 месяцев – 39,5 г. У телят, получавших ежедневно ДПД, соответственно, была выше – в 1,08; 1,04; 1,25 и в 1,02 раза. Из табличных данных видно, что рост массы копытцевой кости до четырёхмесячного возраста развивается одинаково с первой и второй фалангами. Затем до возраста 9 месяцев
резко возрастает и в возрасте 14 месяцев несколько понижается.
Между ростом массы костей III и IV пальца тазовой конечности достоверной разницы нет. Но с возраста телят четыре месяца интенсивность прироста массы костей четвёртого пальца, получавших ДПД, значительно выше. Из данного анализа следует, что ДПД благоприятно влияет не только на развитие вен, но и рост массы костей пальцев и в целом на развитие костной ткани автоподия.
Вопросы для контроля
- Кроме опорной функции, чем является скелет для организма животных?
- Как влияет функция на массу костной ткани, а костная ткань – на функцию?
- Гиперфункция влияет на рост костной массы или нет?
34. Отношение суммарного диаметра вен к массе, прочности костей и к степени ДПД
Вначале выводили коэффициент суммарного диаметра вен, а затем высчитывали количество костной ткани, приходящейся на миллиметр квадратный площади вены. Потом проверяли зависимость его от степени дозированного принудительного движения (ДПД). Было установлено, что отношение количества костной ткани на один миллиметр квадратный диаметра вены зависит не только от ДПД, но и от расположения фаланг в звене пальцев.
У новорожденных телят клеточного содержания для первой фаланги коэффициент составил 0,18 г/мм2 костной ткани. Этот коэффициент оказался выше, чем для второй фаланги. К месячному возрасту коэффициент второй фаланги увеличился на 7,1%, а первой, наоборот, снизился на 11,1%. Коэффициент активно возрастал до четырёхмесячного возраста телят, но к девятимесячному - понижался и вновь увеличивался до 14 месяцев. Как указывал профессор В.И.Фёдоров - на лицо волнообразный рост массы при клеточном содержании.
Молодняк ежедневно получавший ДПД, коэффициент был значительно выше и достигал 0,32 г/мм2, но наибольший коэффициент оказался для копытцевой кости – 0,44 г/мм2. Он также с возрастом животных увеличивается неодинаково, хотя всегда был значительно выше, чем для первой и второй фаланги.
Коэффициент прочности первой фаланги опытной и контрольной групп в месячном возрасте достигал 10 кг/см2 и к четырёхмесячному возрасту постепенно снижался. Но у животных опытной группы он был больше до возраста бычков 9 месяцев, а затем оставался на одном уровне до 14 месяцев с контрольной группой.
Данный коэффициент указывает, что у животных контрольной группы, в связи с гиподинамией, происходит ухудшение гемодинамики и ослабление кровоснабжения в пальцев, что, в свою очередь, приводит к ухудшению биофизических свойств копытцевой кости.
Величина отношения суммарного диаметра вен к массе и прочности костей
акроподия у животных, получавших ежедневно ДПД, по сравнению с телятами клеточного содержания (гиподинамия), подтверждает предположение, что гиподинамия отрицательно влияет на структуру костной ткани, приводит к нарушению гемодинамики в костях, к нарушению биофизических свойств костной ткани, к изменению морфофункционального состояния автоподия.
Вопросы для контроля
- Какая количественная зависимость костной ткани установлена на один мм2 вены при гипердинамии и гиподинамии?
- Как влияет гипофункция на структуру костной ткани, гемодинамику и физиические свйства костной ткани?
Заключение
Проблема морфогенеза сердечнососудистой системы включает большое число неизученных вопросов: от образования ангиобласт – до преобразования их с возрастом в органы кровеносной системы. Вопросы морфогенетического становления сердечнососудистой системы требуют глубокого изучения. Нет ни одной стадии в онтогенезе, на протяжении которой были бы исключены динамические преобразования форм и структуры (В.В.Куприянов, 1975), поэтому исследования, раскрывающие эти преобразования по стадиям онтогенеза, имеют не только теоретическое, но важное практическое значение, знание которых поможет избежать воздействие неблагоприятных факторов на развивающийся организм как в плодном, так и в постнатальном периодах развития.
Изучение сердечнососудистой системы совместно с другими органами и системами всегда относилось к числу важнейших биологических проблем. Комплексный подход исследования позволяет изучить до этого неизвестные факторы биологического развития как отдельных органов, так всего организма. В этом направлении проделана определённая работа (Х.Б. Баймишев, 1984; Б.П.Шевченко, Н.А. Труш, В.А.Рябуха, 2007) и получены интересные данные, раскрывающие многие стороны биологических основ формирования продуктивных качеств животных, позволяющие судить о возрастных и породных различиях строения и развития организма млекопитающих.
Установлено, что в разные периоды жизни животного, в зависимости от
интенсивности роста, развития его органов, систем и частей тела, имеют место характерные отличия в обменных процессах и неодинаковом использовании питательных веществ. Все эти факторы находят отражение в особенностях строения, развития сердечнососудистой системы и в её взаимоотношениях с другими органами и системами животных. У взрослого животного приток крови на периферию организма и отток от неё строго к сердцу строго сбалансирован, а у развивающегося организма (зародыша, предплода, плода, новорожденного, молодняка) чаще наблюдается неравномерность, которая недостаточно изучена. Поэтому, для функциональной оценки этого процесса, большое значение должно придаваться всем звеньям кровеносной системы, но наибольшее – микроциркуляторному руслу, свойственному каждому органу и имеющее разнообразную функцию и сложное строение.
Согласно работам В.В.Куприянова (1969), В.В.Куприянова, Я.Л.Караганова, В.И.Козлова (1975), Б.П.Шевченко (1979; 1999, 2001, 2007) в пятичленном микроциркуляторном русле выделяют звено притока и распределения (артериолы и прекапилляры), обмена (капилляры) и дренажное депонирующее звено (посткапилляры, венулы). Но, несмотря на морфофункциональное единство отделов сердечнососудистой системы и её микроциркуляторного русла чаще исследователи рассматривают, изучают их отдельно, допуская при этом неточности. Такое разобщение единой функциональной системы не создаёт целостного представления как о её строении, так и о их функциональной взаимозависимости, особенно, в онтогенезе.
Причинами повышенного внимания к сердечнососудистой системе, к росту органов в онтогенезе является желание исследователей раскрыть особенности их строения, роста, развития и взаимоотношения. Выяснить закономерности распределения и формирования внутренних сред (гомеостаза), влияния их на здоровье животного, познать потенциальные возможности при различного рода патологических процессах, сопровождающихся нарушениями кровообращения и
др. Впоследствии возникли реальные возможности замены сердца искусственным сердцем или отдельных его структур (клапанов), звеньев кровеносной системы – искусственными сосудами и т.д.
Несмотря на значительные достижения в современной ангиологии и в сосуди-
стой хирургии, многие вопросы морфологии сердечнососудистой системы требуют уточнения, а некоторые остаются ещё не изученными или требуют уточнения. Особенно это касается познания органных, видовых и возрастных различий микроциркуляторного русла, где совершаются основные обменные вопросы. От того, насколько полно мы познаем все эти тонкости сосудисто-тканевых взаимоотношений, во многом будет зависеть успех лечебных и профилактических мероприятий, отсюда получение и сохранность нарождающегося молодняка. Этой проблеме, в какой степени, посвящено настоящее учебное пособие.
Кровеносная система очень тесно связана с органами внутренней секреции. Отсюда большое значение имеет разработка вопросов, связанных с адаптацией организма к неблагоприятным факторам внешней среды, особенно, к антропогенным воздействиям, которым в настоящее время уделяется большое внимание. Анализ исследований последних лет показал, что морфология желез внутренней секреции и функциональные особенности их во многом специфичны и определяются сезонными факторами и человеческой деятельностью, возрастом животных и физиологическим состоянием организма (Г.Н.Кассиль, 1983; И.С.Решетников, 2002, М.Ж. Нурушев, Б.П.Шевченко, М.С.Сеитов, 2011). В научной литературе недостаточно обоснованных данных по органам внутренней секреции домашних животных. Подчёркиваем – домашних животных и, особенно, у пушных зверей, дающих «пушное золото».
Исследования показали, что с возрастом плодов, животных после рождения диаметр аорты, артерий, вен, органов и масса частей тела растёт неравномерно (Б.П.Шевченко, 2001, 2003; М.Ж. Нурушев и др., 2005, 2010). Особенно большим колебаниям в росте подвержена дуга аорты и артериальный проток. Разница в диспропорции роста этих двух участков выравнивается только к 1,5 – месячному возрасту телят после рождения и устраняется с облитерацией протока. До облитерации протока в дуге аорты образуется заужение аорты в виде «горловины», что способствует интенсивному кровотоку из луковицы (синуса) аорты в плечеголовной ствол. Последнее обстоятельство способствует интенсивному кровотоку к голове, богатому питательными веществами и кислородом, что, в конечном счёте, приводит к усиленному развитию передней части тела позднего плода, новорожденного, диспропорция которого по отношению к задней части тела выравнивается к месячному возрасту телят после рождения.
Изучение роста диаметра артерий относительно вен и к костям пальцев показало, что в конце зародышевого периода развития усиленно развиваются артерии, а у предплодов рост их выравнивается (Р.Я. Козаченко, 1987). В первую половину плодного развития опять более интенсивно растут артерии, а во вторую - вены, перед родами и до месячного возраста после рождения – артерии и, начиная с 1,5 месяцев после рождения, вены. Подобные данные получены от плодов белых крыс, северного оленя и крупного рогатого скота. Что даёт право предположить о подобной закономерности развития сосудов для всех плацентарных животных.
Изучение развития кровеносной системы в онтогенезе имеет большое морфофункциональное значение. Оно вскрывает механизм образования жидкостной среды зародыша, предплода, плода, новорожденного, образование гомеостаза и «поля роста», в котором происходит размножение, дифференциация и рост клеток, образование и рост зачатков органов и систем организма, т.е. развитие в целом «нового», а лучше - молодого организма. Нарушение кормления, режима содержания, поения может привести в эти возрастные периоды к патологическому искажению или даже к смерти (Б.П. Токим, 1970; П.Г.Светлов, 1778; Б.П.Шевченко, 1983, 1999), что наблюдается в новорожденном, месячном и трёхмесячном возрастах, совпадающие с развитием сосудов, костной ткани.
Полученные данные совпадают с высказываниями физиологов о том, что организм живёт в «текучей воде» (Ю.Т.Техвер, 1972), т. е. в крови движущейся по сосудам, к этому следует добавить и «развивается» (Б.П.Шевченко,1982). Но формирование среды и гомеостаза в развивающемся организме изучено ещё недостаточно. Это одна из актуальных задач, которая ждёт своего разрешения.
Как показали исследования зародышей, предплодов и ранних плодов, когда продолжается ещё формирование и развитие зачатков органов и организма в
целом, ткани их представляют студнеобразную, полупрозрачную массу, с большим количеством воды. Создание такой среды возможно при условии, когда приток крови по артериям на периферию организма будет преобладать над её оттоком по венам к сердцу – это закон. Только в этом случае возможно создание жидкостной среды организма и «поля роста» для деления клеток, развития зачатков органов и систем организма в целом.
Во второй половине плодного периода развития продолжается качественная и количественная перестройка тканей, органов, начинают интенсивно функциони-
ровать системы плода, особенно опорно-двигательная система, появляются в них значительное количество продуктов метаболизма, которые током крови по венам доставляются и нейтрализуются защитными органами. Это обстоятельство способствует опережающему росту вен.
Опыты показали, если беременному животному давать некачественные корма в зародышевом, предплодном периоде развития, когда интенсивно растут артерии, а вены отстают в росте, они чаще погибают в утробе матери от накопления продуктов интоксикации и метаболизма в их организме. Зародыши рассасываются, а предплоды чаще мумифицируются.
После родов, новорожденный из благоприятной во всех отношениях внутренней (утробной) среды матери, попадает в агрессивную внешнюю среду, которая требует от него повышенное количество качественных питательных веществ и кислорода для дальнейшей жизнедеятельности, развития и роста массы тела. В поисках пищи новорожденный начинает активно двигаться, что приводит к накоплению в тканях, органах движения опять продуктов метаболизма. Последнее обстоятельство приводит к активному росту вен.
Период опережающего роста артерий (новорожденный и телята возраста до одного месяца) самый опасный. Именно в новорожденном, в случае нарушения технологии содержания, кормления чаще погибают телята, т.к. собственная их иммунная система не в состоянии защитить организм от инфекции внешней среды. Иммунная система его начинает вырабатывать тела только на 14–15 сутки после рождения, а до этого срока новорожденный должен жить за счёт иммунных тел, поступивших в его организм из молозива матери. Причём, стенка
12-перстной кишки может через собственные структуры, из собственного просвета пропустить их в кровеносную и лимфатическую систему новорожденного в течение 22 – 26 часов после рождения. Через 22–26 часов этот путь поступления иммунных тел из молозива просвета кишки закрывается. Причём новорожденный должен получать материнское молозиво, а не сборное. Любой специалист в этом случае понимает, если во время новорожденный не получить материнское молозиво и что его ждёт.
В этом возрасте артерии обгоняют в росте вены (до 10 дней), создана благоприятная среда, в которой иммунных тел нет, питательных веществ, кислорода в крови новорожденного достаточно, инфекция попавшая в организм не встречает защиты, начинает активно размножаться, что приводит, в конечном счёте, к смерти новорожденного. При этом следует помнить, что новорожденный должен во время (в течение 30 минут после рождения) получить материнское молозиво, а не сборное, которое может привести к диареи.
У животных в связи с особенностями их экологии (образ жизни, условия обитания, кормления, содержания), обнаружены общие закономерности и видовые, половые различия в росте, развитии, структурной организации и характере адаптивных приспособлений сердечнососудистой и нервной системы.
Функциональная обусловленность архитектоники кровеносных сосудов, сетей, сплетений и кавернозных тел, носящих петлевидный, спиралевидный характер, нервов - сетевидный, расположение интрамуральных образований в венозных сосудах, изучаемых органов, находятся в прямой зависимости от особенностей гемодинамики и связаны с экологическими факторами, в которых обитает животное.
Наиболее существенные признаки ростовой структуры звеньев кровеносной системы животных и её распределения связаны с морфогенетическими факторами и формируются у плодов до рождения. Стабилизационный период динамики морфофункционального становления сосудов и органов, приходится на возраст молодняка 12–18 месяцев постнатального периода развития.
Из роста артерий относительно диаметра вен следует, что в постнатальном периоде развития диаметр вен больше диаметра артерий в 1,8–2,2 раза. Законо-
мерность, приведённую выше, необходимо учитывать при разработке рационов силосного и сенажного типов кормления, для того что бы получить жизнеспособный приплод. При этом, дозированное принудительное движение (ДПД) на расстояние 3,5 - 4 км/ час, благоприятно влияет на гемодинамику копытца, развитие копытного рога, площадь опоры и угол наклона.
Рост диаметра артерий и вен всегда опережает рост массы органов, т.к. для их формирования необходимы питательные, минеральные вещества, кислород, приносимые током крови, что является определяющим в развитии массы органов, систем и всего организма.
Проведённые исследования позволили установить не только архитектонику сосудов стенок костных вен фаланг пальцев, но и выявить изменения венозного русла, массу, прочность костей акроподия под влиянием степени дозированного принудительного движения. Точки выхода вен из костей пальцев, как правило, постоянные во всех периодах онтогенеза и чаще не зависят от ДПД. Из этого следует, что они закладываются за долго до формирования костной ткани в зародышевом периоде, когда на кость ещё не действуют динамические силы. Но вместе с тем число основных истоков, суммарный диаметр, клапанный индекс, толщина стенки вен, а также масса и биофизические свойства костей пальцев под действием ДПД ещё слабо изучены.
Обращает внимание сходство вен I и II фаланг и их отличия от III фаланги, что обусловлено особенностью их строения, биомеханической нагрузкой и разницей биодинамических свойств. На I и II фаланге выявлено четыре зоны выхода костных вен: краевая суставная зона проксимального и дистального эпифизов, дорсальная и плантарная поверхности. Основной зоной выхода вен из копытцевой кости является париетальная и подошвенная поверхности, особенно, вблизи угла кости. Эти данные подтверждаются исследованиями И.В.Хрусталёвой (1972 – 1974), Н.А.Слесаренко (1978).
Анализируя морфологию вен I и II фаланги, обратили внимание, что суммарный диаметр вен, клапанный индекс, толщина венозной стенки больше на
плантарной поверхности, дистальном эпифизе, чем на дорсальной поверхности и проксимальном эпифизе. Считаем, что плантарная поверхность вследствие
натяжения сухожилий сгибателей пальцев в момент опоры о почву, создаёт условия способствующие оттоку крови.. Об этом указывают Н.Н.Лебедев (1976), А.Н.Михайлов (1978).
Анализ показывает, что вены, выйдя из костей, часто анастомизируют с сосудами соседних костей пальцев, образуя единый сосудистый комплекс. Наличие значительного количества анастомозов, лишённых клапанов, даёт основание предполагать отток крови в различных направлениях. Следует отметить, что часть вен париетальной поверхности копытца открывается не только в венозный коллектор венчика, но и в дорсальный венозный анастомоз, расположенный под сухожилием общего разгибателя пальцев. Механизм оттока крови из копытца дан в главе 27.
Вены эмиссарии выходят из фаланг пальцев, как правило, через общие с артериями питательные отверстия. Артерии I и II фаланга, сопровождая вены, охватывают их спмралевидно на всём протяжении. Диаметр артерий и вен этих фаланг одинаковый. На III фаланге калибр артерий в 3 – 4 раза больше, чем у вен и не артерия, а вены уже оплетают их, но не на всём протяжении, а только на выходе из кости. Полагаем, что энергия систолической пульсации артерии способствует продвижению крови по внутрикостным венам.
Толщина стенок вен во всех возрастных группах, по мере поднятия их проксимально, возрастает на 50–75% за счёт увеличения количества мышечных клеток и эластических волокон. Об этом указывают профессора К.С.Кириков (1983) и К.С.Кириков, И.С.Решетников (2009).
Как показывают результаты исследований, уровень ДПД оказывает влияние на строение и толщину стенок вен. Она становится толще при гипердинамии и тоньше - при гиподинамии. Толщина стенок вен также зависит от уровня венозного давления (Б.И.Ткаченко, 1979), в связи с этим можно предположить, что в венах I и II фаланги, содержащихся при гиподинамии, венозное давление понижено, что приводит к застойным явлениям в области автоподия.
Клапанный аппарат вен костей пальцев представлен двухстворчатыми клапанами, встречающиеся неравномерно по всей длине вены. Наиболее постоянно они располагаются у выхода из отверстий эмиссарий и в устье с
магистральными венами. В венах III фаланги обязательно на выходе из костных отверстий, что указывает на их участие в оттоке крови как из кости, так и из сети спинковой поверхности копытца. Величина клапанного индекса вен наибольшая в области второй фаланги, что подтверждает затруднённый отток крови из кости, связанный с большой нагрузкой на кость. Об этом указывают Б.В.Криштофоро-
ва (1976; 1980); Л.Г.Харченко, (1981) и др.
И.В.Хрусталёва (1974) Н.А.Слесаренко, С.С.Кудряшов (1976), Н.А. Слесаренко (1978), исследуя питательные отверстия III фаланги, пришли к заключению, что на копытцевой кости расположены самые крупные питательные отверстия и в большом количестве. Видимо, особые условия и степень биомеханической нагрузки, в которой находится кость, нашли отражение в строении её венозного русла, обеспечивающее своевременный отток крови из копытца.
Полученные данные дают основание считать, что отрицательное влияние гиподинамии выражается в изменении диаметра вен и их клапанного индекса, уменьшением толщины стенок вен, застойными явлениями в области автоподия, затруднённым оттоком крови из копытец. Нарушение гемодинамики приводит к изменению структуры и снижению биофизических свойств костей пальцев и к заболеваниям копытец. Бычки, содержащиеся в условиях гиподинамии (в клетках) в сутки давали привесы по 841 г, а получавшие ДПД 3,5–4,0 км/час – 1023 г, что на 21,6% выше по сравнению с животными, содержащимися в условиях гиподинамии. Эта доза ДПД в условиях промышленных комплексов является профилактической от заболевания конечностей и способствует получению высоких привесов.
Таким образом, введение дозированного принудительного движения в технологию выращивания и откорма бычков в условиях промышленного комплекса обеспечивает в стопе высокую гемодинамику, повышает мясную продуктивность и качество мяса, является профилактикой многих заболевании конечностей.
Алфавитно–предметный указатель
А
Абаксиальный - неосевой
АВА - артерио-венозный анастомоз
Автономная (вегетативная) - регулирует деятельность внутренних нервная система органов и систем. Делится на симпатическую и парасимпатическую части.
Автоподий - Стопа, кисть, делятся на базиоподий (заплюсна, запястье), метаподий (плюсна, пястье) и акроподий (пальцы).
Адаптация - приспособление.
Аксиальный - Осевой.
Ампула - Пузырьковидное расширение просвета вен (сосуда).
Анастомоз - Соединения артерий, вен, лимфатических сосудов. Бывают «конец в конец», «конец в бок», «бок в бок» и др.
Ангиобласт - Сосудистая стволовая клетка.
Ангиомер - Отрезок артерии, вены – от клапана до клапана.
Апофиз - Отросток, бугорок, гребень.
Артерия - Сосуд, идущий от сердца.
Артериола - Артериальный сосуд, диаметром 16 – 22 мкм, имеет среднюю оболочку из одного ряда гладких мышечных клеток, ответвляющий прекапилляры, капилляры.
Артрит - Воспаление суставов.
Б
Базальная мембрана - Основная соединительнотканная пластинка, толщиной до 800 нм, лежит между эндотелием и медией стенки сосуда.
Барорецепторы - Нервные окончания, воспренимающие изменения давления
Биконфлюэнтные - Двухкорневые вены.
Бифуркационные - Деление на две трубки (артерии, трахеи).
В
Вагус - Блуждающий нерв.
Wasa wasorum - Сосуды сосудов.
Вены - Кровеносная жила, сосуды идущие к сердцу.
Венула - Венозный сосуд, диаметром до 100 мкм.
Вентрально - Поверхность, конец напраленные в сторону живота.
Г
Геалиновый хрящ - Суставной хрящ.
Гемоамортизатор - Фыполняющий функцию амортизатора.
Гемодинамика - Движение крови по сосудам.
Гемопоэз - Образование форменных элементов крови.
Гипердинамия - Повышенная нагрузка на органы движения.
Гиподинамия - Пониженная нагрузка на органы движения.
Гломус - Сосудистый клубочек, узелок.
Гомеостаз - Механизмы, обеспечивающие постоянство химической и физико-химической среды и устойчивость организма к внешним факторам
Д
ДПД - Дозированное принудительное движение.
Деструкция - Разрушение, нарушение нормальной структуры органов, тканей.
Диафиз - Тело длинной трубчатой кости.
Диафизарные отверстия - Внутренностные отверстия тела трубчатой кости.
Динамические нагрузки - Физические и механические нагрузки .
Диспропорция - Не соответствие росту органов.
Дистальный - Применим к костям конечностей, означает «нижний конец».
Дистрофия - Нарушение питания тканей, органов и организма в целом. Приводит к перерождению тканей и к смерти.
Дорсальный - Край, поверхность направленная в сторону спины.
З
Зейгоподий - Два луча ноги. К ним относят предплечье и голень.
И
Инжектор - Струйный прибор, насос, приспособление для нагнетания, подачи жидкостей, газов в ёмкости, горелки и т.д.
Инспираторы - Мышцы вдыхатели.
Интеграция - Объединение, восстановление, восполнение.
Интима - Внутренняя оболочка сосуда.
Интрамуральные - Внутристенные, внутриорганные нервы, сосуды.
К
Каверна - Встречается часто в кавернозных телах, обозначает расширение сосуда.
Кавернозное тело - Встречается в половых органах, слизистой оболочке носа, сосках. Это сложное переплетение артерий, вен.
Калибр - Диаметр артерий, вен.
Каудально - Назад, в сторону хвоста направлен край, конец, поверхность.
Каудальная полая вена - Задняя полая вена, открывается в правое предсердие.
Коллатераль - Обходной артериальный, венозный путь.
Коллектор - Накопитель, собиратель в данном случае крови.
Компенсация - Возмещение.
Конгломерат - Округлённая, шаровидная масса клеток крови.
Контрактура - Сужение, сокращение.
Краниально - Край, конец поверхность направленная в сторону головы.
Краниальная полая вена - Передняя полая вена.
Л
Лакуна - Углубление в стенке вены.
М
Медиально - Внутрь, отсюда внутренняя поверхность.
Медия - Средняя оболочка стенки артерии, вены
Мезокард - Стадия развития сердца.
Метаподий - Плюсна, пястье стопы, кисти.
Метафиз - Внутренний конец тела трубчатой кости, покрытый, как правило, апофизом.
Механорецепторы - Концевые окончания нервов, воспренимающие механические раздражения.
Миозит - Воспаление мышц.
Миоциты - Гладкие мышечные клетки.
Моноконфлюэнтные - Однокорневые вены.
Мультиконфлюэнтные - Многокорневые вены
Мультифидные - Множественные вены.
Н
Натальный - Плодный.
Некробактериоз - Копытная болезнь (гниль).
О
Облитерация - Запустение, заростание просвета сосуда.
Онтогенез - Индивидуальное развитие особи.
Остеогенез - Развитие кости.
Остеодистрофия - Разрушение, изменение структуры кости.
Остеомаляция - Размягчение кости.
Остеон - Структурная единица компактного костного вещества. Входит в состав Гаверсовой системы.
Остеопороз - Разрыхление, разрушение кости.
Остеоцит - Зрелая костная клетка.
Отверстия эмиссарии - Отверстия, через которые из кости выходят вены – выпускницы.
П
Пальмаорная поверхность - Ладонная.
Парасимпатическая часть - Висцеральная (внутренностная) часть автономной нервной системы.
Париетальная - Стеновая (спинковая) поверхность.
Патология - Искажение органа от нормы.
Перимизий - Наружная соединительнотканная оболочка мышц
Периоссольные сети - Кровеносные, нервные сети надкостницы
Периостеум - Надкостница.
Плантарная поверхность - Подошвенная.
Платизма - Подкожная, широкая мышца шеи.
Поле роста» - Имеется в виду, площадь, объём пространства для размножения клеток зачатков органов.
Поликонфлюэнтные - Многостволовые, многокорневые.
Полушунт - Длиннее шунта, отдаёт прекапилляры, капилляры, формирующие сегменты сетей.
Постнатальный - После рождения.
Предплодный - Период развития от зародыша до плода, для крупного рогатого скота равен 30 – 60 суткам внутриутробного развития.
Препарирование - Метод приготовления анатомического препарата.
Проксимально - Термин «вверх» применим для трубчатых костей конечностей.
Р
Регидная - Напряжённая по химическому составу жидкостная среда.
Резорбция - Разрыхление, рассасывание костной ткани.
Рецептор - Концевое окончание нервов, воспринимающих раздражения внутренней, внешней среды и передающие раздражения в подкорковые, корковые центры.
С
Сафена - Подкожная артерия, вена задней конечности.
Сегмент - Отрезок артерии, вены от одного клапана до другого.
Серозная оболочка - Выстилает грудную (плевра) и брюшную (брюшина) полости и их органы.
Симпатическая часть - Сосудистая часть автономной (вегетативной) нервной системы.
Синовит - Воспаление синовиальной оболочки капсулы сустава, влагалища и др.
Синусоиды - Расширения мелких вен.
Стабилизация - Устойчивый, постоянный рост …
Стилоподий - Один луч ноги - плечевая, бедренная кости
Статистическая нагрузка - Равновесные нагрузки, приходящиеся на аппарат конечностей во время стояния.
Сфинктер - Суживатель.
Т
Терминальная - Конечная, концевая артерия, вена, дуга.
Терморецепторы - Окончания нервов, воспринимающие изменения температуры внутренней или внешней среды.
Трабекула - Соединительнотканная перегородка.
Трахея - Дыхательное горло.
Трифуркационные - Трёх стволовые артерии, вены и т.д.
Трофика - Питание.
Токсины - Ядовитые вещества.
У
Унитрункальные - Одностволовые.
Ф
Фасция - Бывает поверхностная и глубокая соединительнотканная оболочка.
Филогенез - Изучает историческое развитие органов, систем и в целом организм.
Х
Хеморецепторы - Окончания нервов, воспринимающие изменения химического состава внутренней и внешней среды.
Ш
Шунт - Короткое, прямое соединение артерии ит вены.
Э
Экстерна - Наружная оболочка стенки артерии или вены.
Экспираторы - Мышцы выдыхатели.
Эмбрион - Зародыш.
Эндомизий - Соединительнотканная оболочка, одевающая мышечные волокна, пучки.
Эндотелиоциты - Клетки, выстилающие просвет артерий вен,сердца, лимфатические сосуды.
Эрректильное действие - Выпрямляющее, напрягающее половы органы, соски вымени и др.