4.1 Комплементарность
Комплементарность – явление, когда объединившиеся в генотипе доминантные неаллельные гены формируют новый признак. Это взаимное дополнение действия генов. Например синтез интерферона у человека находится под контролем двух комплементарных доминантных генов, расположенных в разных хромосомах.
Расщепление 9 : 7. Известным примером комплементарного взаимодействия неаллельных генов является наследование окраски цветков душистого горошка (Lathyrus odoratus) при скрещивании двух родительских форм с белыми цветками ААвв и ааВВ. В потомстве F1 (АаВв), а также в F2 (фенотипический класс А_В_) будет появляться новая окраска – пурпурная. При этом в F2 соотношение классов с окрашенными цветками (А_В_) и классов с неокрашенными цветками (А_bb; ааВ_ и ааbb) будет соответствовать формуле 9 : 7. Основными пигментами, определяющими окраску цветков душистого горошка, являются антоцианы.
Расщепление 9 : 6 : 1. Данное расщепление наблюдается при наследовании формы плода у тыквы (Cucurbita pepo) при скрещивании двух родительских организмов сферической формы ааВВ и ААвв. В потомстве F1 все гибриды первого поколения имеют дисковидную форму. В связи с тем, что генотипы ААвв и ааВВ фенотипически не различимы в F2 поколении, то они в сумме дают цифру 6. Дисковидная форма возникает в результате взаимодействия двух доминантных генов (А и В), а удлиненная форма плода – как следствие сочетания их рецессивных аллелей (аавв).
Расщепление 9 : 3 : 4. В том случае, когда фенотип одного из родителей (например, имеющего генотип ааВ_) совпадает с фенотипом рецессивной гомозиготы (ааbb), расщепление в F2 будет 9 : 3 : 4. Рассмотрим наследование трех типов окраски шерсти у мышей: дикой, или рыжевато-серой (агути), черной и белой. Окраска дикого типа зависит от наличия гена, определяющего развитие окраски, а также от гена, обусловливающего распределение пигмента по длине волоса. Каждый волос у мышей агути имеет по длине кольцо желтого пигмента, а в основании и на конце волоска черный пигмент. Такое зональное распределение пигментов и создает окраску агути, свойственную диким грызунам (белка, кролик, морская свинка и др.). У черных мышей отсутствует зональное распределение пигмента – волос по всей длине окрашены равномерно. Белые мыши с красной радужной оболочкой глаз, так называемые альбиносы, лишены пигмента. Окраска шерсти у мышей типа агути доминирует одновременно над черной и белой. При скрещивании черных мышей с белыми все гибриды F1 оказываются агути, а в F2 наблюдается расщепление в отношении 9 агути : 3 черных : 4 белых.
Расщепление 9 : 3 : 3 : 1. Известны и такие случаи, когда каждый из двух комплементарных генов способен проявлять свое действие самостоятельно. Одним из таких примеров является наследование формы гребня у кур. Каждая из доминантных аллелей генов обусловливала развитие гребня определенной формы (гороховидной или розовидной), а взаимодействие этих генов определяло развитие новой формы гребня – ореховидной (рисунок 14).
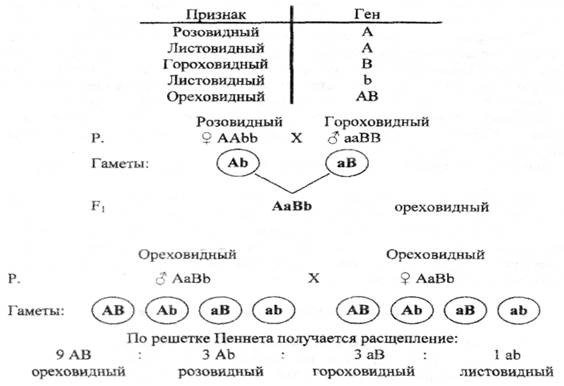
Рисунок 14 – Комплементарное взаимодействие генов