4.2 Эпистаз
Эпистаз и гипостаз – явление подавления действия генов одного аллеля генами другого. Различают эпистаз доминантный и рецессивный. Подавляющий ген называется супрессорным, подавляемый – гипостатическим.
Гены-супрессоры известны у животных (млекопитающих, птиц, насекомых) и у растений. Обычно они обозначаются I или Su в случае доминантного состояния генов и i или su для их рецессивных аллелей (от английских слов inhibitor или supressor). При эпистазе аллель одного из генов подавляет действие аллелей других генов, например А > В или В > А, а > В или b > А и т. д. Эпистатическое взаимодействие генов по своему характеру противоположно комплементарному. В случае комплементарного взаимодействия происходит дополнение одного гена другим. Эпистатическое действие генов по своему характеру очень похоже на явление доминирования, разница состоит лишь в том, что при доминировании аллель подавляет проявление рецессивного аллеля, принадлежащего той же самой аллеломорфной паре. При эпистазе же аллель одного гена подавляет проявление аллеля из другой аллеломорфной пары, т. е. неаллельного гена. Фенотипически эпистаз выражается в отклонении от расщепления, ожидаемого при дигенном наследовании, однако нарушения законов Г. Менделя в этом случае нет, так как распределение аллелей взаимодействующих генов полностью соответствует закону независимого комбинирования признаков.
Доминантный эпистаз (А > В или В > А). Под доминантным эпистазом понимают подавление доминантным аллелем одного гена действия аллельной пары другого гена. Эпистатическая система обнаружена у кур. Некоторые породы кур имеют белое оперение (белый леггорн, белый плимутрок, виандотт и др.), другие же породы имеют окрашенное оперение (австралорп, нью гемпшир, полосатый плимутрок и др.). Белое оперение разных пород кур определяется несколькими различными генами. Так, например, доминантная белая окраска определяется генами CCII (белые леггорны), а рецессивная белая – ccii (белые виандотты). Ген С определяет наличие предшественника пигмента (хромогена), т. е. окрашенность пера, его аллель с – отсутствие хромогена и, следовательно, неокрашенность пера птицы. Ген I является подавителем действия гена С, аллель i не подавляет его действия. В присутствии даже одной дозы гена I в генотипе птицы действие генов окраски не проявится. Поэтому при скрещивании белых леггорнов (CCII) с белыми виндоттами (ccii) гибриды F1 оказываются белыми (СсIi). При скрещивании между собой гибридов F1 во втором поколении имеет место расщепление по окраске в отношении 13 белых: 3 окрашенных (рисунок 15).
При доминантном эпистазе расщепление по фенотипу может выражаться соотношением: 12 : 3 : 1, 13 : 3, 9 : 3 : 4.
|
Признак |
Ген |
|
Окрашенное Белое Супрессор-подавляет Не подавляет |
С с I i |
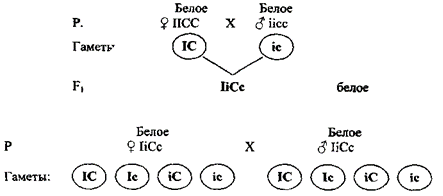
По решетке Пенетта получается расщепление:
9IC : 3Ic : 3iC : 1ic
белое белое окрашенное белое
Суммарное расщепление: 13 : 3
белое окрашенное
Рисунок 15 – Доминантный эпистаз
Рецессивный эпистаз. Под рецессивным эпистазом понимают такой тип взаимодействия, когда рецессивный аллель одного гена, будучи в гомозиготном состоянии, не дает возможности проявиться доминантному или рецессивному аллелям другого гена: аа > В; аа > bb или bb > А; bb > аа.
Примером рецессивного эпистаза является окраска шерсти у собак породы лабрадор. Пигментация шерсти обеспечивается геном В, который в доминантном состоянии дает черную масть, а в рецессивном (b) – коричневую. Имеется также ген С, который в доминантном состоянии не влияет на проявление окраски, но будучи в рецессивном состоянии (сс) подавляет синтез пигмента как черного, так и коричневого. Такие собаки становятся белыми.