Оплодотворение, сингамия, слияние мужской половой клетки (сперматозоид, спермий) с женской (яйцо, яйцеклетка), приводящее к образованию зиготы, которая даёт начало новому организму. У животных оплодотворению предшествует осеменение. В процессе оплодотворения осуществляются активация яйца, объединение гаплоидных наборов хромосом яйца и сперматозоида (амфимиксис), а также, у большинства животных, определение пола развивающегося организма. В результате объединения при оплодотворении отцовских и материнских аллелей возникают новые (в каждом случае уникальные) комбинации наследств, факторов. Таким образом поддерживается генетическое многообразие организмов, которое служит материалом для естественного отбора и эволюции вида. У животных различают два типа оплодотворения – физиологическое моно- и полиспермию. При моноспермии в яйцо проникает один сперматозоид, что обеспечивается особым механизмом блокирования полиспермии. Этот механизм действует на уровне поверхности яйца и яйцевых оболочек; он включает процесс секреции содержимого кортикальных телец и, возможно, какие-то изменения плазмалеммы яйца, а также различные вспомогательные факторы (ограничение поверхности яйца, доступной для сперматозоидов и др.). Этот тип оплодотворения присущ всем животным с наружным осеменением и большинству – с внутренним. У физиологически полиспермных животных в яйцо проникает до нескольских десятков сперматозоидов. Такой тип оплодотворения присущ нескольким группам животных с внутренним осеменением и, по-видимому, возник в эволюции вторично.
Сближение и слияние гамет и их последующие преобразования у животных с разными типами оплодотворения протекают в основном сходно. Чтобы соединиться с яйцом, сперматозоид должен проникнуть через яйцевые оболочки. Для этого у некоторых животных в оболочках яйца имеются отверстия – микропиле. У яиц большинства животных микропиле нет и проникновение сперматозоида в ооплазму происходит с помощью акросомы. Сперматозоид, вступив в контакт с яйцом, осуществляет акросомную реакцию: литические ферменты, заключённые в акросоме, выделяются наружу, вырост акросомы или вся головка сперматозоида проникают через размягчённую ферментами область оболочки и плазмалеммы, гаметы сливаются. С этого момента сперматозоид и яйцо являются единой клеткой – зиготой.
У млекопитающих овулировавшее яйцо, кроме оболочки, окружено несколькими слоями фолликулярных клеток яйценосного бугорка. Сперматозоиды преодолевают этот барьер с помощью содержащегося в акросоме фермента гиалуронидазы, разрушающего желатиноооразную массу, которая связывает фолликулярные клетки между собой. Сперматозоиды млекопитающих становятся способными к осуществлению акросомной реакции лишь после физиологических изменений, называемых капацитацией. Слияние плазмалемм гамет приводит к активации яйца, первым проявлением которой служит кортикальная реакция, обычно сопровождающаяся выделением содержимого кортикальных телец и образованием перивителлинового пространства. Сперматозоид вовлекается в цитоплазму, как правило, целиком; иногда жгутик остаётся снаружи и отбрасывается. Вокруг центросомы сперматозоида появляется лучистость и возникает «семенная звезда», которая позднее, по-видимому, разделяется на две и участвует в формировании веретена первого деления дробления. Перемещаясь в глубь ооплазмы вслед за «семенной звездой», ядро сперматозоида постепенно преобразуется в пузыревидный мужской пронуклеус, его хроматин разрыхляется, а ядерная мембрана распадается на отдельные пузырьки, и мембрана пронуклеуса формируется заново. Эти преобразования происходят одновременно с завершением делений созревания яйца и формированием женского пронуклеуса, который также мигрирует от поверхности в глубь ооплазмы. Оба пронуклеуса сближаются в центре яйца (при равномерном распределении желтка) или в центре области ооплазмы, содержащей меньше желтка, и вступают в контакт. У некоторых животных пронуклеусы сливаются (кариогамия), образуя единое ядро – синкарион, однако обычно они остаются в тесном контакте, не сливаясь, до момента разрушения их оболочек при переходе яйца к дроблению; в этом случае отцовские и материнские хромосомные наборы объединяются на веретене первого деления дробления. Объединение родительских геномов является завершением процесса оплодотворения.
Начальные процессы при оплодотворении
В зрелых половых клетках практически все жизненные процессы подавлены. Клетки фактически находятся в анабиотическом состоянии, и только процесс оплодотворения у большинства видов животных при нормальном развитии является толчком к стре¬мительному выходу яйцеклетки из анабиоза. Существует несколько определений процесса оплодотворения. Оплодотворение представляет собой процесс слияния двух половых клеток (гамет) друг с другом, в результате чего возникает новая особь, генетические потенции которой берут начало от обоих родителей. Согласно другому определению, оплодотворение есть побуждение яйца к развитию, вызванное сперматозоидом, одновременно с передачей яйцу наследственного материала отца. Таким образом, можно говорить о том, что оплодотворение осуществляет две основные функции: половую (комбинирование генов двух родителей) и репродуктивную (создание нового организма), последняя включает инициацию в цитоплазме яйца главным образом тех реакций, которые позволяют продолжать развитие.
Конкретные особенности оплодотворения сильно варьируют у различных видов животных, но происходящие при этом события обычно сводятся к трем основным процессам:
1) процессам, происходящим до соприкосновения спермия и яйцеклетки – так называемое дистантное взаимодействие гамет (этап качественного контроля гамет);
2) процессам, протекающим при непосредственном соприкосновении поверхностей гамет – контактное взаимодействие гамет (этап количественного контроля);
3) процессам, протекающим после вхождения сперматозоида в яйцеклетку – слияние генетического материала спермия и яйца и активация метаболизма яйца для вступления на путь развития.
Дистантное взаимодействие гамет направлено на повышение вероятности встречи сперматозоида с яйцеклеткой. При наружном осеменении многие животные выметывают гаметы непосредственно в окружающую среду – воду. Одновременно туда же могут выметывать половые продукты представители различных видов, поэтому животным необходимы механизмы, препятствующие оплодотворению яйцеклеток одного вида спермиями другого вида. Другая проблема – низкая концентрация спермиев в окружающей среде.
Для решения этих проблем в процессе эволюции сформировались два механизма: видоспецифическое привлечение спермиев и их видоспецифическая активация.
Видоспецифическое привлечение осуществляется посредством хемотаксиса – движения сперматозоидов по градиенту концентрации некоторых специфических веществ, выделяемых яйцеклеткой. Это было доказано для многих животных, в том числе для кишечнополостных, моллюсков, иглокожих и первичнохордовых. Было показано, что яйца кишечнополостного Orthopyxis caliculata не только секретируют хемотаксический фактор, но и способны регулировать время его выделения. Механизмы хемотаксиса раз¬личны.
В 80-х годах XX в. из яиц морского ежа было выделено два специфических хемотаксических агента – пептиды сперакт и резакт. Сперакт, выделенный из яиц Strongylocentrotus purpuratus, состоит из 10 аминокислотных остатков. Резакт, выделенный из студенистой оболочки яиц Arbacia punctulata, содержит 14 аминокислотных остатков. У млекопитающих, когда оплодотворение происходит в верхних отделах яйцевода, основная роль в дистантном вза-имодействии гамет отводится явлению реотаксиса – способности спермиев двигаться против встречного течения жидкости в маточных трубах.
Контактное взаимодействие гамет состоит из нескольких по-следовательных этапов. Первый из них – акросомная реакция. Ее последовательные этапы наиболее полно изучены у морского ежа. Акросомную реакцию сперматозоидов этого вида инициируют сульфатированные полисахариды студенистой оболочки яйца. Они вызывают поступление ионов кальция и натрия в головку спермия и замещение ими ионов калия и водорода. Ионное замещение вызывает целый комплекс внутриклеточных процессов (рисунок 1). Акросомная реакция может быть вызвана и неспецифическим воздействием, например механическим (соударением сперматозоида с любой твердой поверхностью), или при искусственном повышении концентрации ионов кальция в окружающей среде.
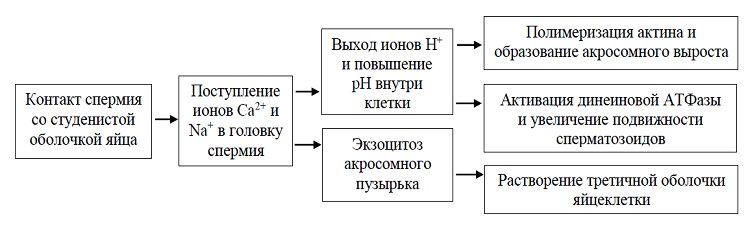
Рисунок 1 – Комплекс внутриклеточных процессов при акросомной реакции
В самом начале акросомной реакции происходит слияние наружной мембраны акросомы с плазматической мембраной сперматозоида и последующий разрыв этих мембран, сопровождающийся экзоцитозом содержимого акросомного пузырька. Изливающиеся из разрушенной акросомы спермиолизины растворяют третичную оболочку яйцеклетки. В результате выхода ионов водорода из головки спермия наступает следующий этап акросомной реакции – образование акросомного выроста. Это осуществляется за счет того, что внутренний участок мембраны акросомы начинает выпячиваться вследствие полимеризации глобулярного актина и образования актиновых микрофиламентов. До активации спермия полимеризация актина, вероятно, блокирована регуляторным белком, а увеличение внутриклеточного рН снимает этот блок.
Акросомный вырост вступает в контакт с наружной поверхностью яйцеклетки – желточной оболочкой. Именно этот момент является решающим для взаимного «видоспецифического» узнавания яйцеклетки и сперматозоида. Это узнавание осуществляется благодаря комплементарному взаимодействию особого белка биндина, встроенного в мембрану акросомного выроста (бывшей внутренней мембраны акросомного пузырька) с соответствующим рецептором на желточной оболочке яйцеклетки. Впервые биндин был выделен из акросомы спермиев Strongylocentrotus purpuratus. Он представляет собой нерастворимый белок с молекулярной массой 30 500 Да, имеющий видоспецифические различия. Проведенные биохимические исследования показали, что биндины близкородственных видов морских ежей различны. В свою очередь из желточных оболочек яиц морских ежей был выделен крупный гликопротеиновый комплекс, который специфическим образом связывал биндин.
Следующий этап процесса оплодотворения – реакция активации метаболизма яйцеклетки. Вслед за реакцией видоспецифического узнавания (образования комплекса между биндином и его рецептором на желточной оболочке яйцеклетки) образуется бугорок оплодотворения. С началом этого процесса можно говорить об активации яйцеклетки. Формирование бугорка оплодотворения сопровождается (как и в случае формирования акросомного выроста) полимеризацией актина. Мембраны верхушки акросомной нити и бугорка оплодотворения сливаются и образуют канал, по которому содержимое сперматозоида поступает в яйцеклетку. Участок мембраны сперматозоида встраивается в мембрану яйцеклетки, где может сохраняться длительное время (у морского ежа иммунохимическими методами этот участок мембраны сперматозоида был обнаружен на стадии личинки).
Дальнейшая реакция активации яйцеклетки связана с функционированием инозитолфосфатной системы, все компоненты которой в неактивном состоянии присутствуют в плазматической мембране яйцеклетки.
Инозитолфосфатная система контролируется белковыми компонентами, локализованными на плазматической мембране яйцеклетки, которые изменяют свою конформацию после взаимо-действия биндина с мембранным рецептором. Такими компонентами являются G-белок и фосфолипаза С. После контакта сперматозоида с плазматической мембраной яйцеклетки в первые секунды независимо от инозитольной системы регуляции резко повышается проницаемость плазматической мембраны яйцеклетки для ионов Na , что в конечном итоге обеспечивает быстрый блок полиспермии (рисунок 2).
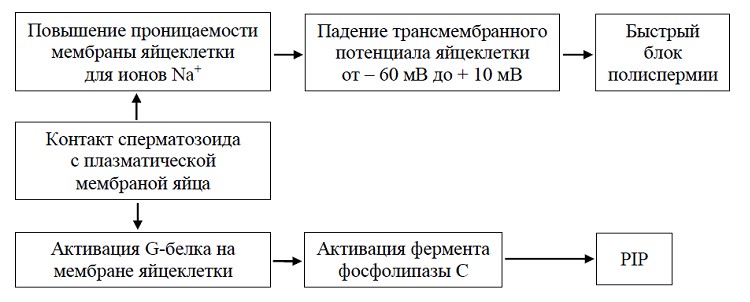
Рисунок 2 – Схема событий, происходящих при контакте сперматозоида с плазматической мембраной яйцеклетки морского ежа в процессе оплодотворения
Основными компонентами инозитолфосфатной системы являются производное углевода инозита – фосфатидилинозитол-4,5-дифосфат (PIP) и фермент фосфолипаза С. Под действием фосфолипазы С фосфатидилинозитол-4,5-дифосфат расщепляется на две составляющие: диацилглицерол (DAG) и инозитолтрифосфат (IР3). Диацилглицерол остается в плазматической мембране яйцеклетки, а инозитолтрифосфат выходит в цитоплазму яйцеклетки. Диацилглицерол с помощью другого мембранного фермента – протеинкиназы С – оказывает влияние на Na/H-обменник, обеспечивая отток ионов Н из цитоплазмы и повышение концентрации ионов Na+ в яйцеклетке. Отток ионов Н+ из цитоплазмы приводит к повышению внутриклеточного рН, что в свою очередь стимулирует ускорение синтеза белка в цитоплазме яйцеклетки и синтез ДНК в мужском и женском ядрах.
В свою очередь инозитолтрифосфат, выходящий из мембраны в цитоплазму яйцеклетки, воздействует на эндоплазматический ретикулум – внутриклеточное «депо» кальция и усиливает высвобождение ионов Са в цитоплазму. Выход Са в цитоплазму активирует целый ряд метаболических реакций. Одна из них заключается в увеличении потребления яйцом кислорода. Интересен тот факт, что такое потребление кислорода практически не связано с митохондриями, которые используют его для синтеза АТФ. Вероятнее всего, кислород используется для окисления таких компонентов яйца, как, например, оболочка оплодотворения. Одновременно окисляются различные липиды цитоплазмы. Кроме того, ионы Са, как и повышение внутриклеточного рН, оказывают влияние на интенсивность синтеза ДНК в мужском и женском ядрах. Другим кальцийзависимым эффектом является активация фермента НАД-киназы, которая превращает НАД в НАДФ, что в конечном итоге обеспечивает синтез новых липидных компонентов плазматической мембраны, необходимых для последующего процесса дробления (рисунок 3).
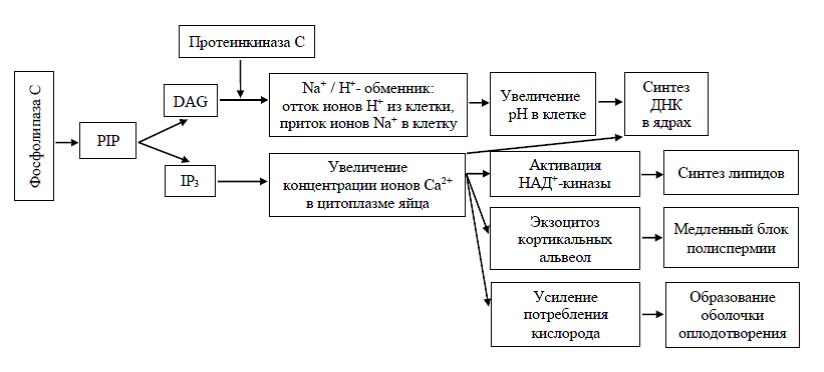
Рисунок 3 – Схема событий, происходящих при последовательной активации фосфолипазы С и протеинкиназы, приводящих к изменению внутриклеточного pH в оплодотворенной яйцеклетке
И наконец, повышение концентрации ионов Са в цитоплазме яйцеклетки приводит к экзоцитозу кортикальных альвеол – кортикальной реакции, которая создает медленный блок полиспермии. Кортикальные альвеолы – это многочисленные пузырьки, содержащиеся в кортикальном слое неоплодотворенной яйцеклетки. При экзоцитозе кортикальных альвеол (кортикальной реакции) из них в пространство между плазматической мембраной яйцеклетки и плотно прилегающей к ней желточной оболочкой высвобождаются следующие вещества:
- вителлиновая деламиназа – протеолитический фермент, разрывающий связи между плазматической мембраной и желточной оболочкой;
- сперморецепторная гидролаза – протеолитический фермент, освобождающий осевшую на желточной оболочке сперму от связей с этой оболочкой;
- гликопротеид, который насасывает воду в пространство между желточной оболочкой и плазматической мембраной, вызывая тем самым их расслоение и образование перивителлинового пространства между ними;
- фактор, способствующий затвердению желточной оболочки, которую теперь называют оболочкой оплодотворения;
- у иглокожих, кроме того, из кортикальных альвеол высвобождается структурный белок гиалин, который участвует в формировании гиалинового слоя, располагающегося под плазматической мембраной.
Таким образом, активация яйцеклетки включает комплекс протекающих в ней реакций, которые затрагивают практически все компоненты яйца.
Поздние реакции оплодотворения включают активацию синтеза ДНК и белкового синтеза. Известно, что еще в период оогеза в самой яйцеклетке запасается большое количество мРНК, до активации находящейся в заблокированном состоянии. В результате комплекса процессов, происходящих при активации яйца (повышение концентрации Са, повышения внутриклеточного рН), происходит деблокирование функции мРНК и начинается интенсивный синтез белка. Такое резкое повышение белкового синтеза начинается уже через несколько минут после проникновения сперматозоида в яйцеклетку. Итак, можно говорить, что интенсивность белкового синтеза не зависит от синтеза новых мРНК в яйцеклетке, а определятся наличием уже запасенных «материнских» мРНК.
Первый и второй блоки мейоза
Полное завершение мейоза без участия сперматозоида, внедряющегося в яйцеклетку, происходит лишь у немногих видов животных (некоторые кишечнополостные, иглокожие – морской еж). У большинства животных он останавливается в конце профазы (как правило, на стадии диплотены-диакинеза). В этом случае говорят о наступлении первого блока мейоза, который у многих позвоночных может продолжаться годами.
Первый блок мейоза снимается во время овуляции. Однако и после этого мейоз все же не доходит до конца – наступает второй блок мейоза (послеовуляторный блок), краткосрочный. Для завершения мейоза необходимо снять этот блок активирующим агентом. В роли такого агента могут выступать сперматозоид, изменение солевого состава окружающей среды, механическое воздействие и т. д.
Всех животных можно поделить на три группы в зависимости от того, как эти остановки мейоза накладываются на процессы овуляции и оплодотворения:
1) мейоз останавливается один раз – на стадии диакинеза и снимается при оплодотворении (губки, некоторые представители плоских, круглых и кольчатых червей, моллюски и т. д.);
2) первый блок мейоза наступает на стадии диплотены-диакинеза профазы мейоза и снимается во время овуляции; второй блок мейоза наступает на метафазе первого деления созревания и снимается при оплодотворении (некоторые губки, немертины, моллюски, практически все насекомые; к такому типу блока мейоза относятся также яйцеклетки млекопитающих – собаки, лисицы и лошади, у которых первый блок мейоза наступает еще в эмбриональный период развития);
3) первый блок мейоза наступает на стадии диплотены-диакинеза профазы мейоза также в эмбриональный период развития и снимается овуляцией; второй блок мейоза наступает на стадии метафазы второго деления созревания, т. е. уже после выделения первого полярного тельца (практически все хордовые; у летучих мышей второй блок мейоза приходится на анафазу второго деления созревания).
Процессы, протекающие после вхождения сперматозоида в яйцеклетку
У большинства животных сперматозоид входит в яйцеклетку целиком, включая хвостовую часть; у некоторых видов жгутик остается на поверхности. Но даже если жгутик с окружающими его митохондриями проникли в яйцеклетку, они не играют никакой роли в процессах дальнейшего развития, а разрушаются в цитоплазме яйцеклетки. Митохондрии передаются зародышу только материнским организмом.
Попав в цитоплазму яйцеклетки, сперматозоид сразу же пово-рачивается шейкой по ходу дальнейшего движения. Хроматин в ядре сперматозоида деспирализуется. Ядерная оболочка распадается на мелкие пузырьки, что делает возможным воздействие цитоплазмы яйца на компактный хроматин спермия.
Белки, благодаря которым хроматин спермия сохраняет конденсированное неактивное состояние, заменяются на белки сходного типа яйцевого происхождения. Эта замена приводит к деконденсации хроматина спермия.
Хроматиновая масса удерживает остатки исходной оболочки сперматического ядра на своей поверхности. Вскоре по периферии этой массы выстраиваются новые мембранные пузырьки, которые соединяются с участками прежней оболочки и образуют новую оболочку мужского ядра. Ядро сперматозоида теперь называют мужским пронуклеусом.
Когда в яйцеклетке завершатся деления мейоза, хроматин ядра также деспирализуется. Женское ядро получает название женского пронуклеуса.
Прежде чем сблизиться, пронуклеусы проделывают сложные движения («танец пронуклеусов»). Сначала мужской пронуклеус движется внутрь яйца перпендикулярно поверхности и независимо от положения женского пронуклеуса. Этот отрезок пути называют «дорожкой проникновения». Затем оба пронуклеуса двигаются навстречу друг другу по «дорожке копуляции».
Движения мужского пронуклеуса осуществляются, по-видимому, благодаря «отталкиванию» микротрубочек, растущих от «мужской» центриоли, которая находится в непосредственной близости к мужскому пронуклеусу, от поверхностного слоя яйцеклетки. У морского ежа процесс сближения пронуклеусов продолжается около 1 ч. На стадии пронуклеусов синтезируется ДНК и удваивается хромосомный набор за счет формирования сестринских хроматид. В конце стадии пронуклеусов начинается конденсация хромосом – идет подготовка к первому делению мейоза. Оболочка пронуклеусов разрушается и конденсированные биваленты занимают свои места на веретене первого деления мейоза. Таким образом, 5-фаза первого клеточного цикла в жизни нового организма приходится на стадию пронуклеусов.
Ооплазматическая сегрегация
Непосредственно после проникновения сперматозоида в яйцеклетку или воздействия активирующего агента начинаются интенсивные перемещения цитоплазмы яйцеклетки. Такие перемещения, а в некоторых случаях и возможные расслоения цитоплазмы, носят название ооплазматической сегрегации. Перемещения играют решающую роль для дальнейшего развития зародыша: в результате ооплазматической сегрегации намечаются основные элементы его пространственной организации.
У различных видов животных ооплазматическая сегрегация протекает по-разному. У некоторых кишечнополостных происходит лишь расслоение цитоплазмы яйцеклетки (ооплазмы) на внешний ободок (эктоплазму) и внутреннюю массу (эндоплазму). Основная масса питательных веществ сосредоточена именно в эндоплазме. У морского ежа, например, до оплодотворения под всей поверхностью яйца рассеян красный пигмент эхинохром. После оплодотворения он концентрируется в виде пояска в экваториальной зоне.
В этих случаях сегрегационные процессы симметричны по меньшей мере относительно полярной оси яйца. Часто сегрегация нарушает полярную симметрию и приводит к выделению в яйцеклетке меридиональной плоскости, которая соответствует сагиттальной плоскости и проходящей в ней дорсовентральной оси будущего зародыша. Примером может быть яйцеклетка амфибий.
Решающим для определения дорсовентральности будущего организма является поворот оплодотворения. Он заключается в повороте в вертикальной плоскости примерно на 30° всего кортикального слоя яйцеклетки относительно внутренней массы желтка. Плоскость поворота совпадает с сагиттальной плоскостью, а направление – с вентродорсальным направлением: та сторона яйцеклетки, с которой частицы кортикального слоя опускаются, становится вентральной, а сторона, на которую они поднимаются, дорсальной.
Поворот оплодотворения контролируется цитоскелетными компонентами – микротрубочками. Применение микротрубочковых ядов (колхицин и т. д.) подавляет поворот оплодотворения. Микротрубочки, которые осуществляют поворот оплодотворения, растут из центриоли, внесенной сперматозоидом. Микротрубочки, достигшие в процессе своего роста вегетативного кортекса, полимеризуются очень активно и загибаются своими внешними концами (плюс-концами) по направлению будущего поворота кортекса. У ряда животных для ооплазматической сегрегации важное значение имеет сократимость актиновых микрофиламентов кор¬тикального слоя яйцеклетки, которая активируется ионами Са.
Контрольные вопросы
1. Дайте определение термину «сингамия».
2. Опишите процессы, происходящие при оплодотверении яйцеклетки сперматозоидом.
3. Какие процессы происходят при акросомной реакцции?
4. Что такое ооплазматическая сегрегация?