13 Оптимизация технологии посева яровой пшеницы в условиях Северо-Востока Казахстана
Глубина посева представляет собою важнейший прием обеспечения высокой (приближенной к лабораторной) полевой всхожести семян, создания присущего сорту начального (стартового) ритма ростовых процессов и темпов дифференциации конуса нарастания, получения здоровых конкурентоспособных равномерных, дружных всходов [44].
Различные сельскохозяйственные культуры в зависимости от биологических особенностей по-разному реагируют на глубину посева семян. В естественных экосистемах осыпающиеся семена прорастают преимущественно с поверхности почвы, находясь под защитой мульчирующего слоя растительных остатков и верхнего биогенного слоя почвы. В агроэкосистемах поверхностный слой почвы пересыхает, и человек взял на себя обязанность создания эффективного ложа для семян. При создании эффективного ложа учитываются потребности семян для прорастания на глубине их посева. К их числу относятся – оптимальная влажность, температура, питательные вещества, доступ кислорода, биологическая способность проростков укореняться и преодолевать верхний слой почвы для выноса конуса нарастания зародышевого побега (точки роста) на ее поверхность.
В последние два десятилетия на примере однодольных злаковых культур (пшеница, ячмень, рожь, рис) показано, что глубина посева в значительной мере зависит от сортовых особенностей семян. Биологическим критерием предельной оптимальной глубины посева сорта служит средняя длина колеоптиле – органа, ответственного за выход проростка на поверхность почвы.
По данным R. E. Allan и O. A. Vogel длина колеоптиле у мягкой пшеницы является полигенным признаком и 10 хромосом несут гены, отвественные за нормальное развитие колеоптиле. Коэффициент наследования этого признака составляет у сортов мягкой пшеницы 82,1 %. Е. П. Алешин и Н. В. Воробьев установили жизненно важную физиологическую роль колеоптиле риса при прорастании семян в условиях затопления. При остром дефиците кислорода колеоптиле участвует в процессах мобилизации запасных веществ эндосперма семени на рост и развитие проростка. При переходе колеоптиле в фазу старения, когда его функции угасают, наблюдаются существенные нарушения в углеводном обмене и происходит массовая гибель семян .
Сорта зерновых культур различаются по длине колеоптиле и их можно по этому признаку разделить на три группы:
- короткоколеоптильные – средняя длина колеоптиле до 4 см;
- среднеколеоптильные – средняя длина колеоптиле до 5 см;
- длинноколеоптильные – средняя длина колептиле 6 см и более.
Условия внешней среды и технология возделывания оказывают определенное влияние на изменение длины колеоптиле, фенотипическая изменчивость которого оказалась значительной, составляя 16–22 %. Это иногда приводит к сдвигу средней длины колеоптиле по сорту в целом. Величину фенотипической изменчивости колеоптиле у сортов целесообразно контролировать по годам и после протравливания семян системными препаратами. Некоторые из них существенно уменьшают длину колеоптиле, требуя корректировки глубины посева и соответствуещей глубины предпосевной подготовки почвы. Однако, несмотря на фенотипическую изменчивость длины колеоптиле, различия между сортами по этому признаку сохраняются, что служит основанием для его использования в качестве оптимального показателя предельной глубины заделки семян.
Скорость продвижения проростка к поверхности почвы под покровом колеоптиле зависит от темпа его роста, который выше у длинноколеоптильных сортов. Так, длинноколеоптильный сорт пшеницы имеет средний темп роста 1,0–2,0 см в сутки и всходит с глубины 3–4 см на 1–2, а сглубины 6–10 см – на 3–4 дня раньше, чем сорта со средним и коротким колеоптиле. Различия в прохождении фенофаз по сортам сохраняются до конца вегетации.
У сортов проявляется четкая реакция на разный уровень глубины заделки семян, прежде всего по их полевой всхожести. С увеличением глубины заделки семян от 4 до 10 см в большей мере снижается полевая всхожесть средне-, а особенно короткоколеоптильных сортов по сравнению с длинноколеоптильными.
Между показателями полевой всхожести семян и интенсивностью развития корневой гнили проявляется обратная зависимость: чем выше полевая всхожесть семян, тем ниже заболеваемость растений корневыми гнилями, и наоборот.
Степень этой зависимости изменяется по сортам и годам. Полевая всхожесть семян снижается при передаче возбудителей через семена в большей мере, чем через почву, а особенно через семена и почву. Вот почему опасно заделывать семена глубже биологически допустимого уровня на почвах, сильно заселенных возбудителями корневой гнили, а также при вынужденном использовании для посева инфицированных семян.
На посевах с глубокой заделкой семян растения в большей мере поражаются корневыми гнилями на протяжении всего вегетационного периода. Самой высокой восприимчивостью к возбудителям болезни отличается эпикотиль. При этом отмечено: чем больше глубина заделки семян, тем длиннее эликотиль и интенсивнее его заражение.
Посев семян глубже средней длины колеоптиле обеспечивает более достоверные различия в восприимчивости к фузариозно-гельминтоспориозным корневым гнилям, чем при заделке на глубину, равную средней длине колеоптиле. Это связано с искусственным нарушением ростовых процессов, прежде всего колеоптиле – органа, ответственного за выход проростка на поверхность почвы.
Образующийся при глубокой заделке семян эпикотиль представляет собою орган – рецептор инфекции, поражение которого нарушает физиологические функции растений в целом [44].
В нарушениях формирования биомассы и сухого вещества на фоне разной глубины заделки семян наблюдается две противоположные тенденции. До фазы кущения при глубокой заделке семян темп ростовых процессов, накопление биомассы и сухого вещества снижается вследствие общего ослабления растений. В дальнейшем вступает в действие компенсационный механизм, обусловленный увеличением плошади питания растений на изреженных посевах. Ростовые и синтетические проиессы усиливаются: увеличивается число придаточных стеблей, вторичных корней, листьев. Однако более интенсивное поражение возбулителями обыкновенной гнили узла кущения и вегегативных органов при глубокой заделке семян тормозит развитие и ослабляет действие компенсационного механизма, что в конечном счете определяет снижение величины урожая зерна.
В литературе распространено ошибочное утверждение, что на глубину залегания узла кущения, а следовательно, и образование узловых корней и побегов положительное влияние оказывает более глубокая заделка семян. На самом деле это не соответствует действительности (рисунок 37)
А. И. Носатовский показал, что основным регулятором глубины залегания узла кущения является свет: узел остается на той глубине, куда проникает свет в период продвижения конуса зародышевого побега. Как только побег подвергается действию света, узел кущения далее не поднимается, а остается на той глубине, какой он достиг при росте в темноте. При изменении глубины посева с 4 до 10 см глубина залегания узла кушения изменяется от 1,9 до 2,7 см. Ближе к поверхности он закладывается в пасмурную погоду, а глубже – в солнечную.
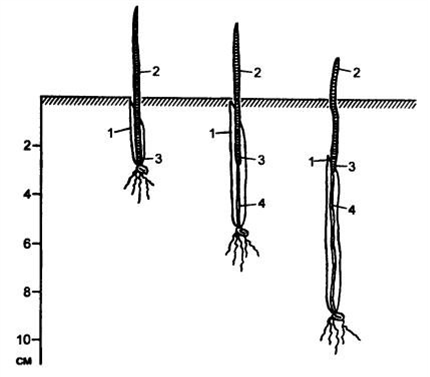 ;
;
1 – колеоптиль; 2 – первый настоящий лист; 3 – узел кущения; 4 – подземное корневидное междоузлие (эпикотиль)
Рисунок 37 – Всходы пшеницы при разной глубине посева семян (по В. А. Кумакову, 1988)
Попытки заглубления узла кущения глубокой зяделкой семян приводят к сниженио энергии кущения, числа и глубины проникновения корней, образованию длинного (более 0,5 см) эпикотиля. Отмеченные компенсаторные механизмы при глубокой заделке семян, обусловленные выпадением растеннй и возросшей площадью питания, не снимают негативных последствий. Вследствие этого при глубокой заделке семян растения испытывают в период засухи двойной стресс: из-за слабого укоренения растений, особенно при рыхлом семенном ложе, и из-за поражения эпикотиля возбудителями корневых гнилей, которое приводит к нарушению координации метаболитических процессов между подземными и надземными вегетативными органами.
Обеспечение нормального, присушего сорту, ритма ростовых процессов при оптимальной заделке семян зерновых культур имеет большое значение в оптимизации фитосанитарного состояния по сорнякам и фитофагам. В первом случае растения приобретают конкурентоспособность, а во втором фитофаги избегают откладывать яйца на быстрорастущие всходы [44]. Кроме того, если и произошло заражение шведской мухой, то ее личинки могут выталкиваться из интенсивно растуших растений – происходит своего рода очишение их от фитофага. Наиболее высокое заражение личинками шведской мухи отмечается на медленно растущих растениях. Активизация ростовых процессов при оптимальной глубине посева и в случае применения других агротехнических приемов (внесении минеральных удобрений) снижает повреждаемость растений шведской мухой в 45 раз.
Среди мероприятий, обеспечивающих оптимальную и равномерную глубину заделки семян, ведущая роль принадлежит предпосевной обработке почвы, задачи которой сводятся к выравниванию поверхности, созданию плотного нижнего и рыхлого верхнего слоя, и благоприятных гидротермических условий для прорастания семян.
Большое значение имеет предпосевное прикатывание, которое хорошо выравнивает, уплотняет почву, уменьшает вертикальную вибрацию сошников и обеспечивает тем самым равномерную заделку сeмян. По нашим данным, всходы на прикатанных участках появляются раньше и дружнее, развиваются интенсивнее, бывают менее восприимчивы к возбудителям корневых гнилей. При этом обеспеченность растений вторичными корнями возрастает на 14,9–20,5 %, урожай зерна повышается на 17,7–23,4 %.
В хозяйствах целесообразно иметь два сорта, различающихся по длине колеоптиле, для обеспечения оптимальной глубины заделки семян в изменяющихся гидротермических условиях внешней срсды. При районировании сортов следует давать им характеристику по длине колеоптиле, что позволит правильнее ориентироваться в глубине заделки семян. При этом при ранних сроках сева целесообразно семена заделывать мельче – на 2–3 см. Это связано с тем, что в слое 2–4 см в весенний период на 2…3,9 °С теплее, чем в слое 6–10 см, а влаги в это время, как правило, достаточно для прорастания семян. При более поздних сроках сева и влаги меньше, и почва уже теплее, поэтому можно сеять глубже: короткоколеоптильные сорта до 4 см, среднеколеоптильные – до 5, а длинноколеоптильные – до 6.
Что касается влияния глубины заделки семян на развитие листо-стеблевых и трансмиссивных инфекций, то оно несушественно или возрастает из-за изреженности посевов при глубокой заделке семян.
Роль оптимизации глубины посева семян на Северо-Востоке Казахстана возрастает в связи с формированием здесь физиологически недозрелых семян со сравнительно слабой дифференциацией зародыша, в том числе колеоптиле, зачаточных листъев, проводящей системы, щитка, эпибласта и других органов. Поэтому зародышевые органы развиваются ослабленными и менее способными пробиваться на поверхностъ почвы через ее толстый слой.
Чем большей физиологической зрелостью обладают семена, тем выше их полевая всхожесть и устойчивость к корневым инфекциям, и наоборот.
Где превалирует физиологическая недозрелость семян зерновых культур, самые густые и дружные всходы появляются при мелкой (2–3 см) заделке семян, если соблюдаются отимальные параметры для эффективного ложа семян:
- семена заделывают во влажный (60 % от полной влагоемкости) слой почвы, содержаший 14–16 мм влаги в слое 0–10 см;
- семена укладывают на твердое (плотность выщелоченного чернозема, например, 1,0–1,1 г/см3) ложе. Для этого глубину предпосевной подготовки почвы проводят на глубину посева;
- семена заделывают на глубину не более средней длины колеоптиле сорта. При этом определяют процент семян, дающих проростки с длиной колеоптиле менее 2,5 см, так как установлена высокая зависимость (r = 0,89 ± 0,09) урожайности зерна яровой пшеницы от процента проростков с длиной колеоптиле более 2,5 см и количества таких проростков с полевой всхожестью семян (r= 0,96 ± 0,08);
- наличие на глубине посева положительной температуры: для набухания семян – минимум 1–2 °С, прорастания и появления всходов – 4–5 °С, оптимальной – 12– 16 °С. При оптимальной влажности почвы сумма эффективных температур (выше 10 °С), необходимая для появления всходов пшеницы с глубины 5 см, составляет 120 °С. При увеличении или уменьшении глубины посева на 1 см сумма эффективных температур повышается или снижается на 10 °С;
- доступ свободного кислорода для гидролиза запасных питательных веществ семени. Энергия, выделяемая при окислении кислородом воздуха сложных органических соединений семени, используется для транспорта метаболитов гидролиза к прорастающему зародышу, а затем к конусу нарастания зародышевого побега;
- наличие растворимых форм питательных веществ, особенно фосфора в почве. Хорошие результаты дает внесение аммофоса или гранулированного суперфосфата в рядки в норме 15–20 кг д. в./га;
- семена прикрывают сверху «пуховым одеялом» с диаметром комков почвы не более 2 см.
Таким образом, глубина посева (посадки) сельскохозяйственных культур, в частности, яровой пшеницы значительно влияет на развитие почвенных и семенных инфекций. При этом по обеим группам инфекций установлена прямая зависимостъ между глубиной посева и их развитием: чем мельче заделка семян, тем ниже развитие болезней, и наоборот.
Глубокая заделка семян сельскохозяйственных культур характерна только для агроэкосистем. Она ведет к нарушению ритма ростовых процессов на начальном (стартовом) этапе развития растений-хозяев, снижает их физиологическую устойчивость, способствуст поражению болезнями, снижает конкурентоспособность к сорнякам и фитофагам. В естественных экосистемах семена растений-хозяев попадают на поверхность почвы и(или) ее верхний слой, покрытый мульчей. В них характерное для агроэкосистем снижение физиологической устойчивости растений к вредным организмам отсутствует. Это служит одной из причин низкого уровня развития болезней в естественных экосистемах.
Вышеуказанные правила создания эффективного ложа для семян и посадочного материала создают предпосылки для повышения полевой всхожести, получения физиологически устойчивых, выносливых и конкурентоспособных посевов, обладающих стартовым ритмом ростовых процессов, начиная с фазы прорастания семян.