Адам денесіндегі бірнеше жүздеген триллион жасушалардың әр қайсысының құрылысы мен қызметі, өзін-өзі реттеу мүмкіншіліктері біркелкі болмайды. Олардың өзара қарым-қатынасы, әрекеттік жүйелердегі ағзалар мен тіндердің байланысы, бір-біріне көрсететін көпжақты әсерлері де әртүрлі. Сондықтан белгілі бір қысқа мерзім милисекунд, минут арасында жасушалар мен тіндерде, жүйелерде сансыз, түрлі-түрлі күрделі үрдістер өтеді. Олардың бір-бірімен қосылуы, жалғасуы және керекті бағытта өтуі организмде ақпараттардың пайда болып таралуына байланысты. Ақпарат жасушаның өзінде немесе сыртқы және ішкі ортаның әсерінен туады. Ақпарат үш түрлі жолмен таралады. Біріншіден, бір жасушадан екінші жасушаға олардың түйіскен жерінен тікелей өтеді. Екіншіден, хабарларды қан мен лимфада және жасушааралық сұйықтықта еріген заттар жеткізеді. Үшіншіден, ақпараттың ең тез таралу жолы электрлік сигнал түрінде яғни жүйке серпіністері (импульстері) арқылы керекті хабарларды бұлжытпай шеткі ағзалардан орталық жүйке жүйесіне одан қайта шеттегі ағзаларға жеткізу.
Тірі организмде тіндерді "қозғыш", "қозбайтын" деп екі топқа бөлуге болады. Қозғыш тіндерге жүйке, бұлшықеттер, сөл бөлетін бездер, ал "қозбайтын" тіндер қатарына дәнекер тін, шеміршек, терінің беткеи тіндер эпителийлері жатады. Қозғыш тін тітіркендірілсе, оған жауап ретінде қозу үрдісі пайда болады, яғни тін тьшьшггық қалпынан қызмет жағдайына көшеді, оның белсенділігі жоғарылайды. Тіннің тітіркендіргіш әсеріне жауап беру қабілеті, оның қозғыштық қасиеті болып саналады. Қозғыштық қозатын гіндердің физиологиялық қасиетінің ең көрнектісі. Қозғыш тіндердің скінші физиологиялық қасиеті қозу үрдісін өткізу. Ет тіндері үшін үшінші қасиет - жиырылу, ал без тіндері қозу салдарынан сөл бөледі. Қозғыш тіндердің физиологиялық қасиеттерін зерттеу үшін көбіне жүйке-ет препараты қолданылады. Ол бақаның шонданай жүйкесімен байланысқан балтыр етінен тұрады.
Тітіркендіру әсерінен қозғыш тінде қозу үрдісі туады. Ол өте күрделі құбылыс. Ол көптеген физиологиялық, биохимиялық және морфологиялық құрылымдық өзгерістермен сипатталады. Тін қозғанда, оның физиологиялық қасиеті - қозғыштығы өзгереді, жасушалардың мембраналық шамасы әрекет потенциалына айналады. Әрекет потенциалы қозудың ең көрнекті белгісі, яғни қозу үрдісінің туғанын әрекет потенциалының пайда болуынан білуге болады. Тін қозған сәтте заттар алмасуы әдеттегіден әлде қандай күшейеді. Осыған орай жылу шығару деңгейі артады, қимыл әрекеттері туады.
Қозатын тіндердің қозғыштығы бірдей емес. Жүйке басқа тіндерге қарағанда қозғыш келеді. Қозғыштық қасиетін салыстырса, қаңқа етінің жүрек етіне қарағанда қозғыштығы жоғарырақ, ал бірыңғай салалы бұлшықеттің қозғыштығы олардан төмен болады. Тіннің қозғыштығы қозу үрдісі кезінде, не қажығанда езгереді.
Қозгыштықтың кезеңдері мен өлшемдері
Тіннің қозғыштығы оны тітіркендіргеннен бастап-ақ өзгереді. Алдымен қозғыштық төмендейді, одан кейін біртіндеп әдеттегі қалпына келеді де, салыстырмалы рефрактерлік (шамалы қасарысу) жоғарылап барып, супернормалды кезең қайта төмендеп, субнормалды кезең қозу үрдісі аяқталарда бұрыңғы тыныштық кезіндегі қалпына келеді. Қозу үрдісі басталысымен тіннің қозғыштық қасиеті құлдырап төмендеп, қаншама әсерлі болса да, біразға дейін келесі тітіркендіргіш күшке жауап бермей қояды. Бұл мезгіл абсолюттік рефрактерлік кезең (нағыз қасарысу) деп аталады.
Қозғыштық дәрежесін анықтау үшін бірнеше өлшем (параметр) қолданылады: қозу табалдырығы, әсердің пайдалы уақыты, хронаксия, лабилдік, аккомодация жылдамдығы. Көбінесе қолданылатын өлшем - қозу табалдырығы. Қозу табалдырығы (реобаза) деп қозу үрдісін тудыратын тітіркендіргіш күшінің ең аз өлшемін айтады. Тін неғұрлым қозғыш болса, оның қозу табалдырығы соғұрлым төмен болады. Тітіркендіргіш ретінде электр тогы қолданылса, табалдырығын рео-база дейді. Әсердің пайдалы уақыты. Тітіркендіргіштің әсері, оның күшіне ғана байланысты емес, әсердің ұзақтығына да байланысты болады.
Хронаксия. Хронаксия деп тінді қоздыратын екі реобазаға тең күш әсерінің ең қысқа уақытын (ОҒ) айтады. Қозғыш тіннің хронаксиясы қысқа болады. Оның мөлшері хронаксиметр аспабымен өлшенеді, мс. есебімен (сигмамен) белгіленеді.
Аккомодация жылдамдыгы. Аккомодация лат. – accomodatio -бейімделу. Белгілі бір тінді, ағзаны қоздыратын токтың әсері оның күшіне, әсер еткен уақытына ғана байланысты емес, токтың үдеу жылдамдығына да байланысты. Егер әсер ететін тоқтың күшін біртіндеп жәй күшейтсе, ол көп жоғарылатылса да тін қозбауы мүмкін. Тоқ күшінің үдеуіне тін бейімделіп үлгереді. Қозғыш тін ток күшінің үдеуіне бейімделгіш келеді. Демек, аккомодация жылдамдығы тіннің қозғыш дәрежесін сипаттай алады.
Лабильдік. Лабильдік (лат. - labilis - құнақылық, тұрақсыздық) -белгілі тіннің қозғыштық дәрежесін көрсететін өлшемдердің бірі. Лабилдік деп тітіркендіру жиілігіне сәйкес 1 секунд ішінде пайда болатын қозудың ең жоғарғы (максималды) әрекеттік ширақтығын айтады. Ол герц өлшемімен белгіленеді. Мәселен, жүйкенің лабилдігі 1000 гц демек, онда жүйке секундына 1200 рет тітіркендірілсе де бұдан пайда болатын қозу саны 1000-нан аспайды. Тін неғұрлым қозғыш болса, оның лабилдігі соғұрлым жоғары болады.
Лабильдік – қозу үрдісінің – ең қарапайым физиологиялық әсерленістің қаншалықты ұзақ екенін көрсетеді. Қозу үрдісі қысқарса, белгілі уақыт ішінде пайда болатын қозу саны көбейеді де тін лабильдігі арта түседі. әдетте, 1 секундта пайда болатын қозу саны тітіркендіру санына немесе күшіне сәйкес болады.
Лабильдік тіннің абсолюттік рефрактерлік кезеңіне байланысты. Абсолюттік кезең ұзақ болса, тін лабильдігі төмен болады, өйткені абсолюттік рефрактерлік кезеңде тін қандай күшпен тітіркендірілсе де жауап бермей қояды. Сондықтан тітіркендіру аралығы абсолюттік рефрактерлік кезеңінен ұзағырақ болуы керек, сонда ғана тін келесі импульске сәйкес қоза алады. Жеке тіннің өзіне ғана тән қозу қабілеті (қозғыштығы), рефрактерлік мерзімі болады. Демек, тіндер лабильдігі де бірдей емес.
Биопотенциалдар (бейне көрініс)
Қозу барысында мембраналық потенциал (МП) әрекет потенциалына (ӘП) айналады. Тірі организмде көптеген потенциалдар (электр тогы) кездеседі: МП, локалды жауап, ӘП, ілеспелі потенциалдары, постинапстық қоздыру және тежеуші потенциалдар. Осылардың ішінде мембраналық потенциал мен әрекет потенциалы бұрынырақ және әлдеқайда толық зерттелген.
Мембраналық потенциал дегеніміз – протоплазма потенциалымен жасушаның сыртқы бетіндегі потенциалдың арасындағы айырмашылық. Мембрананың сыртқы беті әдетте оң, ал шеткі беті теріс зарядталған. Осциллографты пайдаланып оның микроэлектродтарының бірін ет жасушасының (физиологиялық ерітіндіге малынған) ішіне енгізіп, екіншісін сол жасуша мембранасының сыртқы бетіне жақын бекітсе, осциллографтың сәулесі бірден ішкі электродқа қарай жылжиды да протоплазманың теріс зарядын, биоток мөлшерін көрсетеді. Қаңқа еті және кейбір жүйке талшықтарының мембраналық потенциал шамасы 80-90 мВ. Тірі жасушаның қай қайсысының болса да мембраналық потенциалы болады. Жасушаның тіршілігі тоқтаған сәтте мембраналық потенциал жойылады.
Тірі тінде биопотенциалдардың пайда болуы жөнінде мембраналық-иондық, фазалық, заттар алмасу, көп электрлік теориялар бар. Бұлардың ішінде әсіресе мембраналық – иондық теория танымал. Оны алғаш 1902 жылы неміс ғалымы Ю.Бернштейн болжам ретінде ұсынған. Кейін ағылшын ғалымдары А.Хаксли мен А.Ходжкина бұл теорияны тәжірибе жүзінде алынған көптеген деректермен дәлелдеп берді.
Мембраналық – иондық теория бойынша мембраналық потенциалдың пайда болуы протоплазма мен жасушааралық сұйықтықтағы иондық өлшемдестікке байланысты. Осы себепті мембрананың екі беті әртүрлі зарядты. Қалыпты жағдайда жасушалық мембрананың сыртқы беті «оң», ішкі беті «теріс» зарядталған. Жасуша протоплазмасы мен жасушааралық сұйықтықта иондар түрі мен санының ассиметриясын дәлелдейтін мәліметтер жеткілікті. К катионының протоплазмадағы мөлшері жасушааралық сұйықтықтан 30-50 есе артық, ал Na ионы жасушаралық сұйықтықта 8-10, Cl анионы 50 есе көп. Жасуша сыртындағы мембрананың ультра құрылысы электрондық микроскоп арқылы анықталды. Мембрана 3 қабатты белок-липоид молекулаларынан тұратын және онда диаметрі бірнеше нанометр саңылаулар мен микротүтіктердің бар екені дәлелденді. Теория бойынша мембрана иондарды талғамай өткізе бермейді: ол арқылы кейбір иондар тез, екінші бір иондар баяу өтеді не өтпей ұсталып қалады. Мәселен, Na иондары мембрана арқылы тез өтсе, К катионы баяу өтеді, ал аниондардың көбі және белок молекулалары өтпейді. Бұл оның саңылауларының кең-тарлығына ғана байланысты емес, оған микротүтіктің заряды мен одан өтетін иондар зарядының қарым-қатынасы және мембранадан өтетін заттардың липидтерде еру-ерімеу дәрежесі де себепкер. Ионның не молекуланың аумағы мембрана саңылауынан кіші, липидтерде тез еритін болса, заттары саңылау зарядына қарама-қарсы келсе, ондай ион мен молекулалар мембранадан тез өтіп кетеді.
Парабиоз. Жүйкеге ұзақ уақыт күшті химиялық тітіркендіргіштер әсер етсе, оның қозғыштық қасиеті, лабилдігі төмендейді. Оны парабиоз деп атайды. Парабиоз грек сөзі (раrа - жуық, жақын bios- өмір, тіршілік) тіріге жуық хал немесе өмір мен өлім арасындағы жағдай деген мағынада. Н.Е.Введенский ет-жүйке препаратын алып, оның жүйкесінің етке жақын жерін 2% кокаин ерітіндісіне малынған мақтамен орайды, біраздан соң жүйкені еттен алысырақ жерінен токтың жиілігін не күшін біртіндеп жоғарылата тітіркендіріп, бұлшықеттің сіреспе жиырылу сызығын жазады. Бұлшықеттің жиырылу жауабы кокаин тиген жердегі құбылысқа байланысты өзгеретінін байқайды және парабиоздың дамуы мезгілінде үш сатылы өзгерістерді анықтады: теңестіру, парадоксальдық, тежелу. Бірінші - теңестіру кезеңінде тітіркендіргіштің біртіндеп жиілігін не күшін жоғарылатса бәріне беретін жауабы бірдей болады. Екінші - парадоксалды кезеңде әлсіз (сирек) тітіркендіргіштерге беретін бұлшықеттің жауабы күшті (жиі) тітіркендіргіштерге беретін жауабынан жоғары болады. Қалыпты жағдайда бұлшықеттің жауабы тітіркендіргіштің әсеріне тікелей байланысты жоғарылайды. Парабиоз тереңдеген сайын бірінші кезең екіншісіне, ал екінші кезең үшінші кезеңге айналады. Үшінші -тежелу кезеңінде ет тітіркендіргіштерге жауап қайтармайды. Егер парабиоздық әсерді уақытында алып тастап физиологиялық ерітіндімен жүйкені жуса, парабиоз кезеңдері бірін-бірі кері қарай алмастырып жүйке әдеттегі жағдайға келеді.
Парабиоз кезеңдердері альтерация жасайтын әсерлерге жүйкенің лабильдігінің төмендеуімен түсіндіріледі. Лабильдік төмендеген сайын ол жерде жүйкенің абсолюттік рефрактерлік кезеңі ұзара түседі, қозу үрдісі күшейеді. Қозу өзінің жайылу қасиетінен айырылып белгілі бір жерде тұрақталады. Бұл кезде жүйке тітіркендіргіштерге жауап бермейді.
Қорыта келгенде, Н.Е.Введенский жүйке парабиозы тәжірибесінде қозу мен тежелу үрдісінің бірлігін, тежелу күшті және тұрақты қозу екенін, қозу тежелуге немесе керісінше тежелу қозуға алмасқанда парабиоздың үш кезеңінің байқалатынын дәлелдеді. И.П.Павлов кейін бұл мәліметтерді ми қыртысында туатын қозу мен тежелу үрдістерін және фазалық жағдайларды түсіндіруге қолданды.
Бұлшықет қозғыш тіндердің бірі, демек, басқа қозғыш тіндерге тән қасиеттер - қозғыштық, қозуды өткізу бұған да тән. Мұнымен бірге бұлшықеттің ерекше қасиеті - жиырылу. Ет тіннің қозғыштық дәрежесі козу табалдырығы арқылы анықталады. Көлденең жолақты қаңқа еті жүрек етіне қарағанда қозғыштау келеді, яғни оның қозу табалдырығы бәрінен де жоғары болады.
Ет тіні созылғыш (серпімді) келеді. Оның созылғыштығы резеңкеге қарағанда шүбәсіз, яғни босатқанда ет талшығының ұзындығы тура бастапқы созбай тұрғандағы қалпына келеді. Бұлшықеттің негізгі қызметі және физиологиялық қасиеті - жиырылып жазылуы. Жиырылу белгілі бір тітіркендіргіш әсеріне берілетін жауап, яғни ет ұзындығының қысқарып қатаю (тонусының жоғарылауы).
Бұлшықет жиырылған сәтте ет қысқарып, осыған орай, оның тонусы күшейсе, мұны ауксотониялық жиырылу деп, қысқарғанмен тонусы өзгермесе, оны изотониялық деп атайды. Ал, ет қысқармастан тек қатаятын болса, оны изометриялық жиырылу дейді. Белгілі бір тітіркендіргіш жалғыз әсер етсе, оған ет бір рет жиырылып жазылады. Мұны жеке жиырылу деп атайды. Ал етті жиі-жиі тітіркендірсе, сандаған жиырылу бірігіп жинақталады да, ет біраз уақыт сіресіп қалады. Осыған орай ет жиырылуы жеке және сіресіп жиырылуы болып бөлінеді.
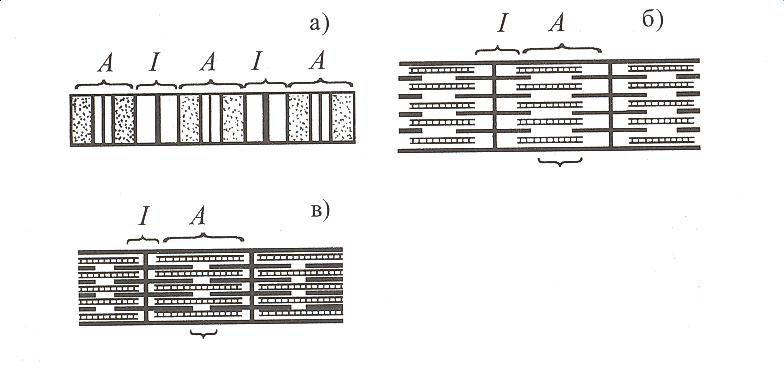
Көлденең жолақты бұлшықеттің жиырылу сырын білу үшін ет жиырылған кезде миофибрилдердің неліктен қысқаратынын, ал солған кезде не себептен ұзарып, қалпына келетінін, сондай-ақ бұлшықеттің қозуы мен қысқаруы арасында қандай тәуелділік бар екенін білу керек. Ол үшін алдымен миоциттің, әсіресе миофибрилдердің құрылымына тоқтай кетейік. Қаңқа еті көлденең жолақты миоциттерден тұрады. Талшық ұзын, цилиндр тәрізді. Оның ұзындығы 10 см-дей, кейде одан да көп, ені 12-70 мкм. Миоцитті қаптаған қабық - сорколемманың саркоплазмаға қарай шыққан көлденең Т - өсінділері болады. Олар миофибрилді саркомераға бөлетін 7. мембраналарының тұсында пайда болады. Сарколемманың іш жағында, миофибрилдер тобының сыртында олармен қатар жатқан саркоплазмалық ретикулум мембрананың Т - өсіндісіне жеткенде кеңіп Са2+ иондарына толы қуыс (цистерна) қүрады. Осы сарколеммалық бір көлденең Т - өсіндісімен оның екі жағындағы екі ретикулум цистернасының түйіскен жері (жасуша ішіндегі үш элементтен түратын триада немесе үштік) - ішкі түйіспе деп аталады. Бұлшықеттің жиырылып қысқаруы мен қайта босауы саркоплазмдағы миофибрилдердің үзындығының өзгеруіне байланысты. Олар талшықтың бір үшынан екінші үшына дейін созылатын қатар-қатар орналасқан жіпшелер будасы (1-сурет, а, б).
А-дискілері (маңгалдары) мен I және H (а) белдеулері көрсетілген. Босаңсыған (б) және жиырылған (в) миофибрилдегі жуан (миозин), жіңішке (актин) жіпшелері.
Миофибрилдерді әрбір 2,5 мкм-ден соң Z мембрана көлденең 2000-дай бөлікке - саркомераға бөледі. Әр саркомераның қақ ортасында 2500-дей жуандығы 10 нм миозин және 2 мембрананың екі жағына тіркелген миозиннен екі есе жіңішке - диаметрі 5 нм актин орналасады. Актиннің ұшы екі жақтағы миозин (белок) талшықтары арасына жартылай кіріп түрады. Миозин жіпшелерідің күңгірт (А) бөлігінде, ал актин ашық (I) бөлігінде болады. Бұл бөліктер миофибрил бойында бірінен соң бірі кезекпен орналасқан. Қатар-қатар жатқан миофибрилдердің күңгірт бөліктері бірыңғай бір қатарда, ашық бөліктері екінші қатарда орналасқандықтан, микроскоппен қарағанда жолақ-жолақ болып көрінеді. Миозин шиыршықты 150 молекулалардан түрады. Әр жіпшенің үшында екі жүмыр (глобула) басы болады. Осы домалақ өсінділері арқылы (көлденең көпірше) миозин актинге жабысады. Миозиннің осы өсіндісінің АТФ қышқылын ыдырататын ферменттік қасиеті бар. Бүл қасиеті актинмен байланысқанда 10 есе жоғарылайды. Актин шиыршықты жіпшелерден түрады, олардың ішінде әрбір 40 нм аралығында домалақ басы бар тропонин молекуласы және тропо-миозин белогі кездеседі. Тропомиозин жіпшелері жиырылмаған етте актиннің миозин өсінділері жабысатын жерін жауып, миозиннің актинге жабысуына кедергі жасайды. Ет жиырылған кезде талшықтар неліктен қысқаратынын түсіндіру үшін қазіргі З.А. Хаскельдің "белок талшықтарының жылжуы" теориясы айрықша қолдау тауып отыр. Бұл теория бойынша ет жиырылған сәтте актин талшықтары миозин бойымен сырғып толығынан олардың ара-арасына кіреді. Мұның салдарынан миофибрилдердің ашық бөлімі (I) қысқарады, тіпті жоғалып та кетеді, ал күңгірт бөлімі (А) оның қақ ортасындағы тек миозин жіпшелерінен тұратын ашыңқы (II) тілімі жоғалып, одан әрі күңгірттене түседі және күңгірт (А) бөлімдері бір-біріне жақындайды (1-сурет, в). Осы сәтте мнофибрилді жай микроскоппен қараса, оның көлденең жолақтығынан айырылғанын көруге болады. Актин және миозин жіпшелері ет жиырылғанда қысқармайды. Актин жіпшелері миозин жіпшелерінің арасына енгендіктен саркомера қысқарады. Бірнеше саркомераның қысқаруы миофибрилді әжептәуір қысқартады. Актиннің миозиндер арасына кіруі миозиндердің көлденең өсінділері, актомиозиннің ферменттік қасиетінің жоғарылауы және саркоплазмада Са2+-иондары деңгейінің көтерілуімен байланыстырылады.
Бірыңғай салалы ет ішкі қуыс ағзалардың қабырғасындағы ет қабағын және кейде топталып қыспақ (сфинктер) құрады. Бұл ет ішек-қарын, ас қорыту бездері түтіктерінің, қан мен лимфа тамырларының, несеп шығаратын мүшелердің, жатырдың, бронхының т. т. қабырғасында кездеседі. Көлденең жолақ етке қарағанда бұл бұлшықеттің организмдегі көлемі аз болады және оның орындайтын қызметі де қалыпты жағдайда онша көзге түспейді (білінбейді). Олай болғанмен потология кезінде, мәселен, асқазан мен он екі елі ішектің түйіскен жеріндегі қыспақ (пилорус) үзақ уақыт жиырылатын болса ас қарында көпке дейін түрып қалады, бара-бара асқазан созылып төмен түсіп кетеді. Мүндай халде тамақты жүтқан сайын ауру қатты қиналатын болады. Мұндай мысалдар көп. Қысқасы, дәрігер үшін бірыңғай салалы бүлшықеттің қасиет ерекшөліктерін жетік білудің маңызы зор. Бірыңғай салалы бұлшықеттің физиологиялық қасиеттерінің негізінде оның морфологиялық ерекшеліктері жатады. Бұл етте барлық жасушаларға тән ультрақүрылымдар бар, тек ретикулумдары аз және жиырылуға қатысатын миофиломенттер (актин мен миозин) саркоплазмада ретсіз қалай болса солай орналасқан. Бұл бұлшықеттің миоциттер мембранасы бұлшықетке қарағанда пиноцитоз құбылысына икемірек болады және кальций каналдары басқа да екі валентті иондарды (Ва2+, Зг2*), Л/а+ ионын өткізеді. Кальций каналдары изоптин дәрісімен жабылып қалады. Мүның әсері медицинада қолданылады. Бірыңғай салалы ет қозған сәтте АҮФ комплексі ете жай ыдырайды. Бұлшықеттің протоплазмасында Л/а+, СГ иондарының мөлшері көлденең жолақ етке қарағанда жоғарырақ болады және миоцит мембранасы арқылы иондардың алмасуы тез өтеді. Осыған байланысты МП әсіресе автоматиялық қасиеті бар салалы еттерде түрақсыз 30-70 мв-тай, ал ӘП-ы МП-дан 10-20 мв қана артық болады. МП мен ӘП-ның мөлшері каңқа етінікінен төмен. Бұлшықетінде әр миоцитпен жүйке талшығы түйіспе қүратын болса, бірыңғай салалы бүлшықеттің әр миоцитімен жүйке талшығы түйіспе қүрмайды. Бірақ тін қозған кезде жүйкетелген миоциттер ғана емес жүйкетелмеген миоциттер де қозады. Өйткені салалы бұлшықеттің миоциттері бір-бірімен нексус арқылы байланысқан. Нексустерден әрекет потенциалы бір жасушадан екінші жасушаға сол күйінде өте береді. Сондықтан бірыңғай салалы ет функциялық синцитийлерге жатады.
Бірыңғай салалы еттің қаңқа еті секілді қозғыштық, қозуды өткізу, жиырылу қасиеттері болады. Мұнымен бірге ол ерекше пластикалық және автоматиялық қасиеттерге де ие. Бірыңғай салалы еттің көлденең жолақ етке қарағанда қозғыштық қасиеті төмен, хронаксиясы, қозу табалдырығы жоғары, абсолюттік рефрактерлік кезеңі ұзақ, лабилдігі төмен болады.
Бірыңғай салалы бұлшықеттің жекелеген жиырылуы өте ұзаққа созылады. Әсіресе латенттік және жазылу (босаңсу) кезеңі қаңқа етінен көп ұзақ. Мысалы, қоянның ішек етінің латенттік кезеңі 0,25-1 с, асқазан етінің жеке жиырылуы 5 с, ал құрбақаның қарын етінің жекеленген жиырылуы бір минут, балтыр етінің жеке жиырылуы 0,11-0,12 с, латенттік кезеңі 0,01 с.
Осыған орай салалы бұлшықеттің сіреспе жиырылуы (тетанус) минутіне 10-12 рет тітіркендіргенде болады. Қозу үрдісі бірыңғай салалы ет арқылы өте баяу таралады (2-10 см/с). Мұның себебі бұл етте пайда болған ӘП-ның жылдамдығы тінге әсер еткен тітіркендіргіштің күшіне және тітіркендіргенде бірден қозатын ет талшығының санына байланысты.Бірыңғай салалы етті қатты созса, ол келесі тітіркендіруге дейін ширықпай сол ұзындығын сақтайды. Мұны дәлелдеу үшін бақа асқазанының бір ұшын миографтың үстіңгі жағына бекітіп, екінші ұшына белгілі бір салмақты (гирді) іліп созса, гирді алып тастаған соң ет сол созылған күйінде келесі тітіркендіруге дейін қалады. Ал көлденең жолақ балтыр еті гирді ала сала қайтадан бұрынғы қалпына келеді. Кейбір бірыңғай салалы еттердің (асқазанның, ішектің, несеп өткізгіш түтіктің) автоматиялық қасиеті болады. Олар тітіркендіргіштің әсерінсіз-ақ өздігінен белгілі бір ырғақпен жиырылады және ол түрлі сырттан келген әсерлер арқылы жиілейді, кейде тежелуі де мүмкін. Бұл бұлшықеттің автоматиялық қасиетінің туу себебінің бірі - оның МП-ның тұрақсыздығы. Бірыңғай салалы етке гуморалдық заттар - гормондар, медиаторлар, иондар әсер етіп қозғыштық қасиетін өзгертеді, қозу үрдісімен қатар тежелу де туғызады.Симпатикалық және парасимпатикалық жүйке жүйелері бірыңғай салалы еттердің жиырылуын, тонусын қарама-қарсы өзгертеді (біреуі жиыратын болса, екіншісі сол етті босатады).