4 Морфология и ультраструктурная организация прокариот
4.1 Морфологическое разнообразие прокариот
Мельчайшие одноклеточные организмы растительной природы и не содержащие хлорофилл относятся к бактериям. Они получили название грибов-дробянок за биологические особенности и способность размножаться простым поперечным делением. Относятся к классу Schizomycetes порядку Eubacteriales.
Бактерии имеют величину от десятых долей микрометра (1 мкм=0,001 мм) до 10–15 мкм в длину и от 0,2 до 1,0 мкм в диаметре. Также есть организмы значительно больших и меньших размеров. При стабильных условиях размеры и формы бактерий могут быть постоянными свойственных данному типу, роду или семейству это результат процесса длительной эволюции их. Но в других случаях под влиянием условий существования, например, температуры, осмотического давления или антибиотиков размеры и форма бактерий может изменяться. Размеры бактерий выражают в микрометрах (мкм). Один микрометр равен 1000 нм (нанометров). Так: 1мм = 103мкм = 106нм = 10А = 109пм (пикометров). Отдельные компоненты бактерий измеряются также в нанометрах. Структуру микроорганизмов изучают с помощью электронной микроскопии целых клеток и их ультратонких срезов.
Бактериальная клетка является живым организмом, которому присуща сложная внутренняя организация, которая включает морфологически и биохимически обособленные структуры, выполняющие определенную жизненную функцию.
По таким морфологическим признакам бактериальных клеток как форма, величина, расположение и сочетание клеток осуществляется классификация бактерий.
Бактерии по форме подразделяются на три основные группы:
1) шаровидные (кокки);
2) палочковидные (бактерии, бациллы, клостридии);
3) извитые (вибрионы, спириллы и спирохеты).
Кокки (Coccus) это бактерии имеющие шаровидную форму с диаметром в 1–2 мкм. Встречаются кокки и других, например, бобовидной, сферической, ланцетовидной форм. В зависимости от направления деления кокков образуются своеобразные формы. Основой классификации на роды является расположение кокков в культуре. При делении кокков в одной плоскости и при быстром расхождении дочерних клеток образуются моно или микрококки (род Micrococcus). Если при делении в одной плоскости дочерние клетки остаются скрепленными попарно, то образуются диплококки (род Diplococcus) или цепочки, состоящие из нескольких соединенных клеток – стрептококки (род Streptococcus). Деление, происходящее в двух взаимно перпендикулярных плоскостях, при котором кокки соединяются по 4 клетки образует – тетракокки (род Tetracoccus). При делении клетки в трех взаимно перпендикулярных направлениях кокки образуют форму пакетов или тюков, которые состоят из 8, 16 и более клеток называются сарцины (род Sarcina).
При беспорядочном делении кокков с образованием кучек похожих на гроздья винограда образуются формы, носящие название стафилококки (род Staphylococcus) рисунок 3. В этой группе микроорганизмов встречаются виды, вызывающие болезни человека и животных.

1 – моно или микрококки (род Micrococcus), 2 – диплококки (род Diplococcus), 3 – стрептококки (род Streptococcus),4 – тетракокки (род Tetracoccus), 5 – сарцины (род Sarcina),6 – стафилококки (род Staphylococcus)
Рисунок 3 – Родовая принадлежность кокков в зависимости от расположения клеток после деления
Палочковидные микроорганизмы бывают цилиндрической формы с обрезанными, заостренными или закругленными концами. Формы, которые не образуют споры называются бактериями (род Bacterium). Палочковидные бактерии, имеющие разветвленную форму относятся к коринебактериям (род Corinebacterium). Палочковидные бактерии сильно варьируют по величине. Есть бактерии, которые в оптическом микроскопе (0,1х0,15 мкм) стоят на грани видимости и есть в несколько десятков микрон как серобактерии. Размеры палочковидных бактерий бывают в пределах в длину 1–5 мкм, и имеют диаметр 0,5–1 мкм. При неблагоприятных условиях в цитоплазме некоторых палочковидных бактерий образуется – спора, представляющая собой тельце круглой или овальной формы.
Спорообразующие микроорганизмы относятся к бациллам (род Bacillus) или называются клостридиями (род Clostridium). Палочковидные бактерии могут располагаться в пространстве беспорядочно, попарно (диплобактерии, диплобациллы). Это самая многочисленная и разнообразная группа микроорганизмов, среди них есть сапрофитные и патогенные виды.
Извитые бактерии к ним относятся бактерии, имеющие один или несколько завитков. Вибрионом (род Vibrio) называется бактерия, имеющая один неполный завиток спирали в виде запятой. Спириллой (род Spirillum) называется бактерия, которая имеет несколько спиралевидных завитков. Если микроб имеет большое количество мелких завитков, то есть штопорообразно извитую форму то тогда будет называться спирохетой (порядок Spirochaetales) (рисунок 4). Морфологию и внутреннюю структуру бактерий изучают на препаратах живых и неживых культур.
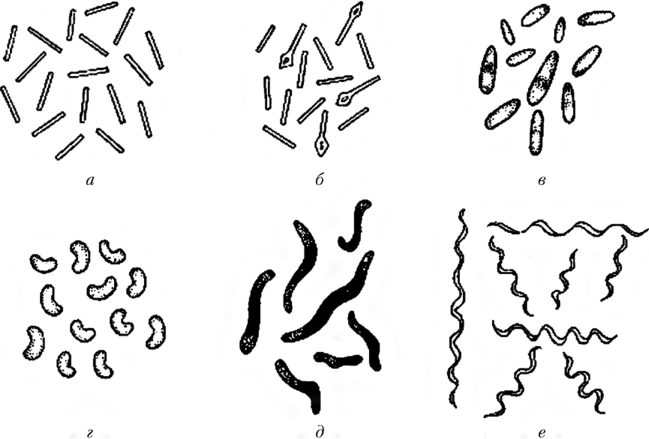
а – бактерия (род Bacterium), б – коринебактерия род Corinebacterium), в – бацилла (род Bacillus), клостридия род Clostridium), г – вибрион (род Vibrio) д – спирилла (род Spirillum), е – спирохетта (порядок Spirochaetales)
Рисунок 4 – Палочковидные и извитые формы бактерий
4.2 Поверхностные структуры прокариот
Бактериальная клетка состоит из поверхностных структур и протопласта, к поверхностным структурам относятся: клеточная стенка, капсула, слизистый чехол, жгутики и ворсинки они располагаются снаружи цитоплазматической мембраны (ЦПМ). Протопласт это цитоплазматическая мембрана (ЦПМ) вместе с цитоплазмой.
Клеточная стенка это многослойное образование, плотно покрывающее цитоплазматическую мембрану. Она является важным и обязательным структурным элементом бактериальной клетки на её долю приходится от 5 до 50 процентов сухого вещества клетки. Клеточная стенка содержит специфические полимерные комплексы, не содержащиеся в других структурах клетки.
Составу химических веществ клеточной стенки и строению свойственно постоянство, что являются одними из важных диагностических признаков. В клеточной стенке большинства бактерий различают три слоя: наружный – липопротеиновый, средний – липополисахаридный и внутренний – мукопептидный (муреиновый). Внутрений слой обеспечивает жесткость (ригидность) клеточной стенки. В зависимости от строения стенки прокариоты окрашиваются по-разному и делятся на грамположительные и грамотрицательные. Способ окраски был предложен в 1884 г. датским ученым Х. Грамом.
Во внутреннем муреиновом слое бактериальной клетки в значительном количестве содержатся мукополипептиды, полифосфатнуклеотиды и липиды. Именно этот муреиновый слой наиболее выражен у грамположительных бактерий. Этот слой у грамотрицательных бактерий бывает незначителен. Сверху он прикрыт более мощным липополисахаридным и липопротеидным слоями.
Протопласты, сферопласты и L-формы бактерий, их свойства
При воздействии на грамположительные бактерии литическими ферментами, лизоцимом и эндопептидазами получаются клетки, лишенные клеточной стенки – протопласты. Они всегда сферической формы в них осуществляются основные процессы жизнедеятельности такие как дыхание, синтез белков, нуклеиновых кислот, спорообразование. Протопласты могут увеличиваться в размерах, фиксировать азот, и они не способны ресинтезировать клеточную стенку. Они редко делятся, так как рецепторы фагов локализованы в клеточной стенке протопласты не адсорбируют фаги.
В старых культурах в условиях несбалансированного роста, под влиянием иммунных сывороток обнаруживают бактериальные клетки частично лишенных клеточной стенки – сферопласты. Их также можно получить под влиянием пенициллина в гипертоническом растворе сахарозы или NaCL (осмотические стабилизаторы). В виду частичного сохранения клеточной стенки сферопласты способны адсорбировать фаги, размножаться и при устранении факторов, вызвавших их образование, могут легко реверсировать в исходную клеточную форму. Большие размеры, отсутствие клеточных мембран типа мезосом, высокая чувствительность к осмотическим условиям являются общими свойствами протопластов и сферопластов.
При антибиотикотерапии или при нарушении биосинтеза пептидогликана в стенке бактериальной клетки приводящее к частичному или полному лишению его образуются L-формы бактерий. У них нарушена функция размножения, но сохранена функция роста вследствие чего увеличиваются до гигантских размеры клеток (до 50 мкм). Образуются сильно вакуолизированные шаровидные, нитевидные, грушевидные формы. L-формы метаболически активны, способны к делению и слиянию элементов, растут в виде характерных колоний с врастанием в питательную среду пигментированным центром и кружевным краем (в виде яичницы). L-формы болезнетворных бактерий патогенны. Заболевания, обусловленные реверсией L-форм, протекают длительно с меньшей смертностью, но с большей инвалидностью. Образование этих форм бактерий имеют приспособительное значение для переживания неблагоприятных условий.
Функции клеточной стенки бактериальных клеток:
1) формообразующая функция;
2) защищает от воздействия факторов окружающей среды;
3) защищает от внутреннего осмотического давления;
4) участвует в транспорте необходимых клетке веществ и ионов;
5) является барьером для проникновения антибиотиков и токсических веществ;
6) изолирует содержимое клетки от гидролитических ферментов;
7) содержит транспортные белки и гидролитические ферменты;
8) содержит специфические рецепторы и антигены;
9) создает условия для межклеточного взаимодействия при конъюгации, а также между патогенными бактериями и тканями высших организмов [4].
4.3 Слизистые поверхностные структуры: слизистые слои, капсулы и чехлы. Химическая природа и функции слизистых поверхностных структур
Клеточная стенка у большинства прокариот бывает окружена веществом слизистой природы, которые в зависимости от структурных особенностей называются капсула, слизистый слой или чехол. Образованных при биосинтезе клеткой органических полимеров.
Слизистое образование аморфного строения, которое обволакивает клетку, сохраняя связь с клеточной стенкой называется капсула. Микрокапсулой называется капсула, если её толщина меньше 0,2 мкм, толщина макрокапсул больше 0,2 мкм. При микроскопировании световым микроскопом с контрастным окрашиванием препарата макрокапсулу бывает видно. На наличие капсулы оказывает влияние условий культивирования и принадлежность к определенным штаммам бактерий. При мутации бактерии могут терять способность образовывать капсулы образуя бескапсульные формы. Капсула служит средством защиты бактерий от неблагоприятных воздействий окружающей среды, например, высыхания, также от защитных факторов организма таких как действие антител или фагоцитоз, также защищает от химических веществ. Если капсула образуется вокруг целой группы клеток, которые соединены между собой как, например, Azotobacter chroococcum то есть окружены единой слизистой капсулой то их называют зооглеями.
Образованию капсулы у сапрофитных микробов способствует избыток в питательной среде углеводов и недостаток азотистой пищи или низкая температура культивирования для некоторых видов.
Чехлики некоторых нитчатых бактерий образуются в результате затвердевания наружных слоев слизи, чехлы в отличие от капсул имеют несколько слоев с разным строением. При метаболизме бактерий с окислением восстановленных соединений металлов в чехликах отлагается окислы, например, у железобактерий отлагается гидроокись железа и клетки приобретают особую прочность.
У большинства капсул прокариот основным химическим компонентом являются гомо-или гетерополисахариды.
Функционально капсулы и чехлики защищают клетку от механических повреждений, высыхания, создают дополнительный осмотический барьер, препятствуют проникновению фагов, могут служить источником запасных питательных веществ, осуществляют связь между соседними клетками и способствуют прикреплению клеток к различным поверхностям.
4.4 Жгутики, пили (фимбрии); типы строение и разнообразие функций
У некоторых видов бактерий имеются органы передвижения, называемые жгутиками. Это очень тонкие (0,01–0,05 мкм), длинные винтообразно скрученные пучки фибрилл. Число, размеры и расположение их являются постоянными для определенных видов.
По расположению жгутиков подвижные бактерии делятся на четыре группы: монотрихи – бактерии с одним расположенным на конце жгутиком; лофотрихи бактериальные клетки, имеющие пучок жгутиков на одном конце; амфитрихи имеют по одному или несколько жгутиков на обоих полюсах; перитрихи имеют множество жгутиков по всей поверхности клетки (рисунок 5).
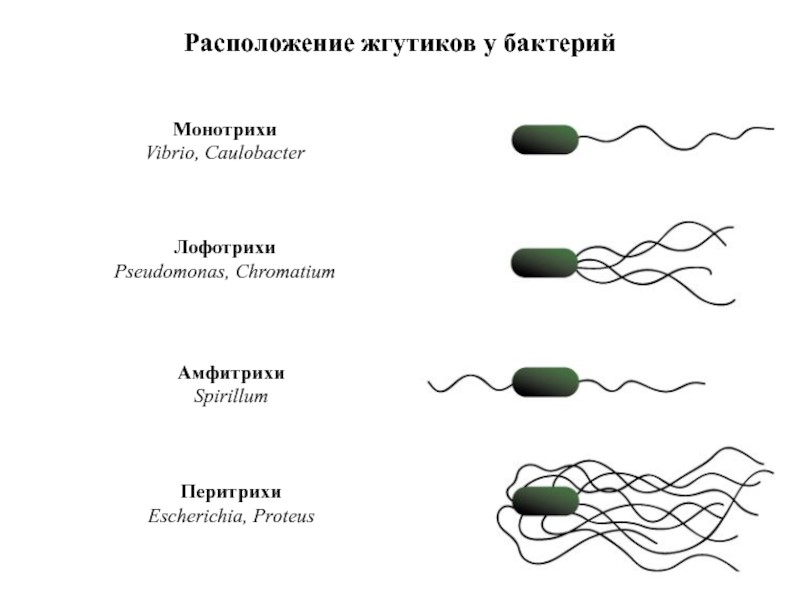
Рисунок 5 – Расположение жгутиков у бактерий
Жгутики состоят из фибриллярного белка флагелина по своим антигенным свойствам отличающегося от белков протоплазмы. Морфологически жгутик состоит из трех частей это спиральная нить, «крюк» часть, находящаяся вблизи поверхности клетки и базальное тельце. Крюк обеспечивает гибкое соединение нити с базальным тельцем и состоит из белка, отличающегося от флагелина – фибриллярного белка жгутиков [4].
Характер движения бактерий определяет спиралевидное или волнообразное движение жгутиков, происходящих из-за ритмических сокращений фибрилл. Движение монотрих происходит всегда по прямой линии с совершением иногда воронкообразных движений на одном месте. Перетрихи совершают беспорядочные кувыркательные движения. Внешние факторы определяют направление активного перемещения подвижных бактерий. Движение бактерий относительно источника химического вещества называется хемотаксисом. Химические вещества в этом плане делятся на две группы: инертные и вызывающие таксисы (эффекторы). Эффекторы могут быть аттрактантами (привлекающими веществами) и репеллентами (вещества, отпугивающие бактерий). К аттрактантам могут относиться аминокислоты, витамины, нуклеотиды, сахара. Репелленты – спирты, фенолы, неорганические ионы также аминокислоты и другие. Аэротаксис также является проявлением хемотакса. Движение к свету свойственный фототрофным бактериям называется фототаксис. Способность бактерий перемещаться по силовым линиям магнитного поля Земли или магнита – магнитотаксис. В клетках бактерий совершающих магнитотаксис найдены частицы, заполненные железом в форме магнетита (Fe3O4), они выполняют функцию магнитной стрелки и носят название магнитосомы. Движение бактерий перемещаться в направлении увеличения или уменьшения вязкости раствора называется вискозитаксис.
Реснички (фимбрии, пили, ворсинки), расположены по всей бактериальной клетке. Не имеют отношения к движению и обнаруживаются как у подвижных, так и неподвижных форм бактерий. Это состоящие из белка пилина прямые, тонкие и короткие белковые цилиндры, отходящие от поверхности клетки. Тоньше жгутиков длина их 0,2–2,0 мкм, диаметр 5–140 нм. Располагаются по перетрихиальному типу или по полюсам. Изучены два типа фимбрий общего типа и половые. Фимбрии общего типа обеспечивают бактериальной клетке гидрофобное свойство и адсорбции бактерий на почвенных частицах, растительных и животных клетках. Половые ворсинки, или F-пили способствуют склеиванию бактерий друг с другом при половом процессе. F-пили в качестве конъюгационного тоннеля обеспечивает передачу ДНК от клетки донора реципиенту. Фимбрии относятся к необязательным клеточным структурам.
4.5 Цитоплазматическая мембрана, особенности ее состава, структуры и функций у бактерий
Цитоплазматическая мембрана (ЦПМ) отделяет содержимое клетки от её стенки. Она является обязательным структурным элементом любой клетки, и нарушение целостности ЦПМ приводит к гибели. ЦПМ составляет от 8–15 % сухого вещества клеток, у большинства прокариотных клеток является единственной мембраной. Цитоплазматическая мембрана бактерий это биологическая мембраной, состоящая из белков и липидов, иногда обнаруживается значительное количество углеводов. Функция липидов ЦПМ – поддержание механической стабильности и обеспечение гидрофобности.
Белки цитоплазматической мембраны являются ферментами. По аминокислотному составу они отличаются от других клеточных белков.
Цитоплазматическая мембрана выполняет следующие функции:
1) барьерную функцию;
2) в ЦПМ локализованы ферменты, поэтому одной из функций является ферментативная;
3) в ЦПМ также расположены переносчики цепи электронного транспорта, поэтому она выполняет энергетическую функцию;
4) участвует в репликации хромосомы;
5) объединяет клетку, то есть интегрирует в единое целое;
6) транспортную функцию (используются разные механизмы мембранного транспорта).
Существует 4 типа транспортных систем: активный транспорт, перенос химически модифицированных молекул, пассивная диффузия и облегченная диффузия.
4.6 Внутрицитоплазматические мембранные структуры бактерий
Глубокие впячивания в цитоплазму и разрастания цитоплазматической мембраны способствуют возникновению внутриплазматической мембраны. Среди них различают:
1) локальные впячивания ЦПМ различающиеся формой, размерами и расположением в клетке называются – мезосомы;
2) у фотосинтезирующих бактерий имеются фотосинтетические мембраны (тилакоиды). У зеленых бактерий пигменты содержатся в хлоросомах у цианобактерий в фикобилисомах;
3) основной фермент фиксации СО2 в цикле Кальвина рибулозодифосфаткарбоксилазу содержат карбоксисомы;
4) у водных бактерий, некоторых почвенных бактерий и бактерий обитателей илов газовые вакуоли или аэросомы регулируют плавучесть;
5) магнитосомы – как говорилось выше обнаруживаются в клетках бактерий, которые обладают магнитотаксисом, магнитосомы содержат частицы Fe3O4 [4, с. 22].
4.7 Цитоплазма прокариотной клетки, рибосомы
Содержимое клетки, окруженное ЦПМ, называется цитоплазмой. Состоит из цитозоля (полужидкой коллоидной массы) содержащую ферментные белки, продукты и субстраты метаболических реакций, и набор растворимых РНК. К другой части цитоплазмы относятся разнообразные структурные элементы: внутрицитоплазматические мембраны, рибосомы, генетический аппарат и включения.
Рибосомы бактерий это сложные образования, которые состоят из различных молекул РНК и белков. Они являются местом, где синтезируется белок. Их размеры составляют 15–20 нм. Количество в зависимости от интенсивности белкового синтеза варьирует от 5000 до 90000. Это рибонуклеиновые частицы, состоящие из двух субчастиц с коэффициентом седиментации 30S и 50S. Рибосомы прокариот обладают константой седиментации 70S. Рибосомальные белки выполняют структурную функцию.
Агрегаты, состоящие из рибосом, молекул информационной и транспортных РНК осуществляют синтез белка называются полирибосомами или полисомами. Могут быть связанными с мембранными структурами или находиться в цитоплазме.
4.8 Запасные вещества
Веществами запаса бактерий являются полисахариды (крахмал, гранулеза, гликоген), липиды, полипептиды, волютин, отложения серы.
В клетках некоторых серобактерий были найдены известковые тельца (карбонат кальция), существует предположение, что они выполняют функцию нейтрализации среды, соединяясь с серной кислотой, которая образуется при окислении внутриклеточной серы.
Параспоральными включениями называют кристаллоподобные включения вблизи спор. Такие включения имеет, например, Bacillus thuringiensis грамположительные почвенные бактерии. Эти включения имеют белковую природу и токсичны для насекомых, но в то же время совершенно безвредны для позвоночных животных и растений. Этот вид бактерий широко используют как инсектицид в сельском хозяйстве.
Запасные вещества прокариот – это высокомолекулярные полимерные молекулы, которые бывают иногда отграничены от цитоплазмы белковой мембраной и находятся в осмотически неактивном состоянии. Нахождение их в таком состоянии важно, так как сосредоточение в цитоплазме множества молекул осмотически активных веществ может оказать отрицательное действие на клетку [4, с. 22].
4.9 Генетический аппарат прокариот. Нуклеоид бактерий, строение, размеры, функции и методы его выявления
ДНК (нуклеоид) прокариот – это компактное образование, занимает центральную область в цитоплазме и не отделенное от неё мембраной. У прокариот одна молекула ДНК содержит всю генетическую информацию, носит название бактериальной хромосомы и имеет форму ковалентно замкнутого кольца. В развернутом виде длина молекулы может составлять более 1 мм.
Репликация ДНК происходит по полуконсервативному механизму. Она начинается в точке прикрепления кольцевой хромосомы к цитоплазматической мембране, где находится ферментативный аппарат, отвечающий за процесс репликации.
Соединение ДНК с цитоплазматической мембраной осуществляется с участием мезосом. Процесс репликации идет в двух противоположных направлениях. Появляющиеся дочерние хромосомы остаются прикрепленными к мембране. Параллельно с репликацией молекул ДНК происходит синтез мембраны в области контакта ДНК с цитоплазматической мембраной. В результате происходит разделение дочерних молекул ДНК и оформляются обособленные хромосомы.
В клетках бактерий кроме хромосом часто находятся плазмиды – тоже замкнутые в кольцо ДНК они также способны к независимой репликации. Плазмиды неотличимы от хромосом, содержат дополнительные гены, которые необходимы лишь в специфических условиях. В плазмидах кодируются механизмы устойчивости к антибиотикам, механизмы разрушения специфических веществ и т.д. Также в плазмидах находятся nif-гены необходимые для азаотфиксации. Сохранение плазмид в дочерних клетках обеспечивают специальные механизмы распределения.
Транспозоны это мобильные сегменты в ДНК бактерий, которые могут перемещаться из одной части хромосомы к другой или в ДНК находящиеся вне хромосомы, а также в другие клетки. Мобильные сегменты ДНК неспособны к автономной репликации [4, с. 22].
4.10 Деление бактерий. Разнообразие способов деление
Для бактерий характерно поперечное бинарное деление, при котором образуются две одинаковые дочерние клетки. У грамположительных деление происходит септой то есть путем синтеза поперечной перегородки, идущего от периферии к центру, у грамотрицательных – перетяжкой. Другими типами размножения бактерий являются – почкование и множественное деление. Почкование происходит с помощью дополнительных выростов, при этом выделяется материнская и дочерняя клетки, почкование можно рассматривать как неравновеликое бинарное деление. Множественное деление отмечается у некоторых цианобактерий, при таком делении образуется множество мелких иногда подвижных баецитов.
Бинарное деление происходит в одной или нескольких плоскостях. При делении в одной плоскости клетки не расходятся, это приводит к образованию цепочек палочковидных или сферических клеток. При делении в нескольких плоскостях образуются клеточные скопления разной формы [4, с. 22].
4.11 Типы дифференцировки бактерий
Бактериальная клетка имеет повышенную выживаемость, поэтому ей присуща морфологическая дифференцировка вегетативных клеток. Специализированные клетки отличаются от вегетативных клеток бактерий, способность их формирования запрограммирована в генетическом аппарате. В процессе развития бактерий под действием внешних факторов формируются структуры, относящиеся к категории покоящихся форм. Их назначение обеспечить сохранение вида в течение длительного времени в неблагоприятных условиях. При наступлении или попадания в благоприятные условия покоящиеся формы прорастая, дают начало вегетативным клеткам.
Есть также морфологически дифференцированные клетки, предназначающиеся для размножения. Это баециты и гормогонии цианобактерий.
Морфологически дифференцированные клетки со специальными метаболическими функциями гетероцисты цианобактерий, бактероиды клубеньковых бактерий выполняют фиксацию молекулярного азота атмосферы.
Споры у бактерий являются своеобразной покоящейся формой, основная функция её сохранение наследственной информации в неблагоприятных условиях, в отличие от спор грибов не являются способом размножения. Споры бывают овальными, круглыми или эллиптическими, для усиления устойчивости к механическим воздействиям у некоторых видов бывают «рёбра жесткости».
Особый тип покоящихся клеток грамположительных бактерий, которые формируются в цитоплазме материнской клетки называются эндоспоры. Одна бактериальная клетка формирует одну эндоспору. У эндоспор имеется многослойный белковый покров, наружный и внутренний мембраны и самый толстый слой оболочки – кортекс. У некоторых бактерий образуется двух–трехслойное желатинообразное покрытие аналогичное капсуле бактерий состоящее из липопротеинов и углеводов называемое экзоспориумом. Эндоспоры обладают устойчивостью к высоким температурам и радиации, которые летальны для вегетативных клеток.
Если у спорообразующих аэробных бактерий, образуется спора диаметром не превышающая диаметр клетки, бактерии называются бациллами. Спорообразующие анаэробные бактерии, у которых размер споры превышает диаметр клетки, и они, поэтому принимают форму веретена, называются клостридиями (от лат. clostridium – веретено).
Гифы большинства актиномицетов делятся перегородками на участки, каждый из которых представляет собой будущую экзоспору. Экзоспоры формируются снаружи микробной клетки. Формирование их происходит одновременно с уплотнением и утолщением клеточной стенки. Отличаются ещё от эндоспор отсутствием кортекса и экзоспориума. У экзоспор отсутствует дипиколиновая кислота. Споры актиномицетов являются одновременно покоящимися клетками и репродуктивными структурами. Экзоспоры могут формироваться путём отпочкования от одного из полюсов материнской клетки как у бактерий из рода Methylosinus и Rhodomicrobium.
Встречающиеся у разных групп бактерий цисты морфологически могут не отличаться от вегетативных клеток. Цисты также являются покоящейся формой существования для пережидания неблагоприятных условий среды. Стенки цист состоят из одного или нескольких внешних (экзина) и внутренних (интина) слоев одинакового или разного химического состава и также различных по структуре. В цитоплазме цист мало воды, обмен энергии и веществ сильно замедлен.
Покоящиеся клетки некоторых цианобактерий называются – акинеты. В отличие от покоящихся клеток многих видов микроорганизмов они крупнее вегетативных клеток. Продолговатой или сферической формы, имеют гранулированное содержимое и толстую оболочку. Могут прорастать сразу после образования или после переноса в свежую питательную среду.
Цисты и акинеты образуются в условиях, когда устойчивость к физическим воздействиям у вегетативных клеток нарушается. Они более устойчивее вегетативных клеток, например, к нагреванию и высушиванию [4, с. 22].
Задания для самостоятельной работы
- Группы организмов, относящиеся к микроорганизмам.
- Особенности строения прокариотической и эукариотической клетки.
- Особенности морфологии бактерий. Физиология бактерий.
- Строение клеточной стенки Грам-положительных и Грам-отрицательных бактерий. Окраска по Граму.
- Протопласты, сферопласты и L формы бактерий. Причины рецидивов болезней.
- Строение и функции цитоплазматической мембраны бактерий. Производные цитоплазматической мембраны. Функция цитоплазмы клетки.
- Включения цитоплазмы бактериальной клетки.
- Внешние структурные образования прокариотической клетки, (капсула, эндоспора, жгутики, фибрии, пили), их функции.
- Морфологическое разнообразие бактерий.
- Основные структуры бактериальной клетки.
- Дополнительные структуры бактериальной клетки.
- Особенности строения и функции клеточной стенки бактерий.
- Нуклеоид, его строение и функции.
- Отличия между жгутиками, фимбриями и пилями бактерий.